

" וּמִכָּל-הָחַי מִכָּל-בָּשָׂר שְׁנַיִם מִכֹּל תָּבִיא אֶל-הַתֵּבָה לְהַחֲיֹת אִתָּךְ: זָכָר וּנְקֵבָה יִהְיוּ" (בראשית ה',פסוק יט)
אבי שמידע – המחלקה לאבולוציה, סיסטמטיקה והתנהגות והמרכז לרציונליות, האוניברסיטה העברית ירושלים, גבעת רם .avi.shmida@gmail.com
עמוס קריינר – בית-ספר התיכון "גלילי" כפר-סבא ועמית במחלקה לפיזיקה מכון וויצמן למדע – amos.kreiner@gmail.com
חמוטל קריינר – המעבדה לקוגניציה לשונית, מדעי ההתנהגות, המרכז האקדמי רופין – hamutalk@ruppin.ac.
תקציר: מכיוון שבכל מחזור רבייה של אוכלוסייה בטבע כל נקבה מתרבה רק פעם אחת ואילו כל זכר יכול להפרות נקבות רבות, נראה לכאורה שהמצב היעיל הוא מעט זכרים והרבה נקבות בכל אוכלוסייה. אולם בניגוד לכך, יחס הזוויגים באוכלוסיות של האדם ושל מרבית המינים בטבע הוא 50:50, כלומר מספר שווה של זכרים ונקבות. במאמר זה אנו מסבירים, באמצעות דוגמאות מספריות, מדוע יש מספר שווה של זכרים ונקבות, מצב הנראה לכאורה כלא מיטבי וכשווי-משקל בלתי-יעיל עבור אוכלוסיות של מינים ביולוגיים בטבע. ההסבר מתבסס על העיקרון הדרווינסטי של הברירה הטבעית, על עיקרון הרציונליות של תורת-המשחקים ועל תורות כלכליות. באמצעות כלל ההשקעה השווה, הנגזר מהתיאוריה על ההקצאה הזוויגית של צ'רנוב, אנו נחשפים לעקרון פישר על יחס הזוויגים המיושם לדוגמאות רבות בטבע. המאמר מוליך אותנו מהסבר של יחס הזוויגים, הנראה לכאורה קל להבנה, להסבר שלתופעות מפתיעות בטבע כגון: יחס זוויגים גמיש בצמחים, שכיחות לא הגיונית של זכרים בטבע ובזבוז עצום של גרגירי אבקה בצמחים מואבקי רוח.
===================
מבוא
ברוב היצורים בעולם – צמחים, בעלי-חיים, פטריות וחד-תאיים – היחס המספרי בין הזכרים לבין הנקבות הוא "חצי – חצי", דהיינו מספר הצאצאים הנולדים מתאי זרע של כל זכר יחיד באוכלוסייה שווה למספר הצאצאים המעמידה נקבה[1](Maynard-Smith 1982, Charnov 1982). להלן נכנה יחס זוויגים זה בשם "50:50" כלומר 50% מהצאצאים הנולדים באוכלוסייה הם זכרים ו 50% הן נקבות [2].
לכאורה יחס זוויגים שווה בין זכרים לנקבות נראה טבעי. המדען הראשון שבדק וחקר זאת היה רופא הכתר של מלכת בריטניה, ג'ון אבותנוט[3] John Abouthnot)). כבר בשנת 1710 הוא טען שיחס הזוויגים השווה בבני-אדם, נובע מ"ההשגחה העליונה, כדי שכל גבר ימצא לו אישה באותה ארץ ובגיל מתאים", כלומר כדי שלא יישארו פרטים לא מזווגים בחברה האנושית. למעשה התוצאות שמצא ג'ון אבותנוט יותר מורכבות: בבדיקת התפלגות הלידות שנספרו בעיר לונדון במשך 82 שנה נמצא יחס קרוב ל-50:50 בין מספר לידות הבנים והבנות, עם מעט יותר לידות של זכרים. לטענת אבותנוט, סטייה זו "מאוזנת על ידי ההשגחה העליונה באמצעות הגדלת תמותת הזכרים". כלומר מכיוון שסיכוני התמותה של גברים גדולים מאלה של הבנות, דואגת ההשגחה העליונה שייוולדו יותר גברים כדי למלא את החסר[4]. אבותנוט ממשיך את טיעונו "הנוצרי" כי בשמיים נקבע מספר שווה של גברים ונשים בכל מדינה ואומר: "מכאן נובע שפוליגמיה היא מנוגדת לחוק הטבע ומנוגדת לצדק, וגם מגבילה את ההתרבות של המין האנושי! כיוון שמספר הנשים והגברים שווה, אם גבר לוקח לו 20 נשים, 19 גברים נידונו לחיי פרישות ו-20 נשים לא תופרינה באותה יעילות על ידי גבר אחד!". בעוד אנו מסתכלים ב"חיוך" להסבריו של אבותנוט הרי שמורה לו זכות ראשונים לגבי אבחנת העובדות ובהצגת השאלה – מדוע יחס הזכרים והנקבות שווה לרוב באוכלוסיות בני-אדם ?


התמר המצוי (משמאל)הוא עץ דו-ביתי ואכן באוכלוסיות הטבעיות שלו בקניוני מואב ואדום יחס הזוויגים הוא חתי-חצי באוכלוסיות שנספרו בכל ואדי בנפרד. מאידך רוב מיני משפחת הדקליים בעולם הםבעלי טיפוס מיניות "חד-ביתי חד-מיני" כמו הדקל הנראה מימין, סיאגרוס רומנזוף ("דקל הקוקוס") השכיח בגינון בישראל; בראש אותו גזע רואים מימין תפרחת זכרית שופעת באנטזיס ומשמאל תפרחת נקבית דלת פרחים לאחר ההפריה. צלם א.ש.


אם מסכמים מאות מחקרים שנעשו על יחס הבנים והבנות שנולדו בארצות שונות, מוצאים כי באופן כללי יחס הזוויגים הוא אכן 50: 50 עם סטיות קלות של עד 5%. המחקר המודרני מוצא כי אחוז הפלת עוברי זכרים גדול יותר מאשר עוברי נקבות אולם נולדים רק מעט יותר זכרים מאשר נקבות[5]. יש החושבים כי הסטייה נובעת ממשך-חיים ארוך יותר של הנקבות ויש המסבירים כי תמותה גבוהה יותר של עוברי זכרים נובעת מהצטברות פגמים בכרומוזום Y הזכרי כיוון שזה לא עובר רקומבינציה (אודי מרגליות, מידע בעל פה). אולם יש לציין כי משך חיי הנקבות נמשך כיום הרבה מעבר לגיל הפריון שלהן.
בשנות השלושים של המאה הקודמת עבדה בבריטניה קבוצה של ביולוגים אשר הניחה את היסודות לאבולוציה ולאקולוגיה המודרנית. בראשם היו הסטטיסטיקאי והביולוג רונלד פישר ועמיתו ג'ון הלדיין[6]. הלדיין ידוע בכך שטבע את המונח "מחיר הסקס", כלומר כל אורגניזם שנוקט באסטרטגיה של רביה מינית מפסיד מחצית מיכולתו האבולוציונית ליצירת צאצאים הזהים לו גנטית (תיבה 1), אך היה זה פישר שנתן את ההסבר הרציונלי הראשון והמשכנע מדוע יחס הזוויגים בטבע ברוב היצורים הוא דווקא 50:50. תחילה נסביר מדוע רביה זוויגית היא מאוד לא יעילה לעומת רביה אל-מינית ולאחר מכן ניתן דוגמא מספרית מדוע מושג זה של שווי משקל אבולוציוני 50:50 הוא מאוד בלתי-יעיל. בטבע ברוב היצורים הוא דווקא 50:50. תחילה נעמוד על ההבדלים בין רביה זוויגית לרבייה אל-מינית ולאחר מכן ניתן דוגמא מספרית איך נוצר יחס הזוויגים בטבע, מדוע הוא מהווה שווי משקל אבולוציוני ומדוע יחס זוויגים "50:50" הוא מאוד בלתי-יעיל.
| תיבה 1
מדוע יחס 50:50 מהווה נטל על אוכלוסיית מין ביולוגי, איננו יחס מיטבי (למי?) ויש בו בזבוז עצום ? במינים ביולוגיים רבים של בעלי חיים וצמחים, חלק ניכר מהזכרים כלל איננו מעמיד צאצאים. בבעלי-חיים רבים יש זכר דומיננטי המשתלט על קבוצת נקבות, או שמרבית הנקבות מעדיפות להזדווג עם אותו זכר מועדף. במקרים אלה כל שאר הזכרים כמעט שאינם מזדווגים. זאת כאשר הנקבות נושאות את מרבית הנטל של גידול הצאצאים. ביונקים יש הריון ממושך ולאחריו הנקה מתישה וגידול הילוד; בעופות הנקבות מייצרות ביצים, נושאות אותן ולפעמים גם דוגרות ומגדלות את הצאצאים לבדן. לעומת זאת, במרבית המקרים הזכרים תורמים רק תאי זרע קטנים. מכיוון שכמחצית החומר התורשתי של הצאצא (מחצית מספר הכרומוזומים) מקורה בביצית האם ומחציתה מקורה בתא הזרע שתרם האב, מרוויחים גם הזכרים, למרות ההשקעה הקטנה שלהם, כמחצית הכשרות הדרוויניסטית של כל ילוד.
בעלי חיים עם שני טיפוסי זוויג (חיות חד-זוויגיות). שימו לב להבדלי הצורה בין שני הזוויגים.
|




הצגה סכמטית של רבייה אל-מינית (מימין) ורבייה זוויגית (משמאל). מקור: Scientific American
מדוע רביה זוויגית (בתהליך מיני) אינה יעילה בהשוואה לרבייה אל-מינית? במינים ביולוגיים בעלי רבייה אל-מינית, מספיק שכל פרט באוכלוסייה ייצר צאצא אחד בממוצע במשך כל ימי חייו, כדי שגודל האוכלוסייה יישאר יציב. ואולם אם כל פרט במין בעל רביה מינית, שבו כל נקבה מייצרת רק צאצא אחד, האוכלוסייה בדור הבא תקטן למחצית גודלה של אוכלוסיית ההורים. דוגמא פשוטה זו מדגימה כי אוכלוסייה המתרבה בדרך אל-מינית תגבר בקלות על אוכלוסייה המתרבה בדרך מינית, כאשר כל שאר התכונות ההתנהגותיות והמורפולוגיות זהות. כלומר, באוכלוסייה המינית על הנקבות לייצר פי שניים צאצאים כדי להשתוות להצלחה של האוכלוסייה האל-מינית. לנחיתות לכאורה זו של הרבייה המינית בהשוואה לרבייה האל-מינית בייצור צאצאים, קרא הלדיין "The cost of sex" (Haldane, 1940). הערך האדפטיבי של הרבייה המינית מוסבר היטב במאמרים וספרים רבים (Maynard-Smith and Szathmary,1995; Charnov, 1982; Ridley, 1996 ) ועל כך נדון במאמר אחר (שמידע וכהן, כתב-יד, שמידע וכהן, 1983, שמידע, 1984).
בדוגמא שלהלן ננסה להסביר מדוע כאשר במין הביולוגי ישנם פרטים בעלי שני טיפוסי סקס נפרדים (זכרים ונקבות), כמו בתמר, בחרוב ובמרבית בעלי החיים, יחס הזוויגים הוא בדרך כלל 50:50. למרות שהדוגמא נראית לכאורה פשוטה ובהירה, היא טומנת בחובה רבדים עמוקים של "קונפליקט טובת המין לעומת טובת הפרט", "קונפליקט הורים צאצאים" וקונפליקטים נוספים בנושא האלטרואיזם.
כיצד מתקבל יחס זוויגים שוויוני של 50:50 בין זכרים לנקבות באוכלוסייה ?
או במילים אחרות: כיצד לחשוב נכון מבחינה אבולוציונית על תהליכי הטבע ?
ניקח לדוגמא את אוכלוסיית הברבור השחור המקננת באיי חמורבי בים הלבן. באוכלוסייה הדמיונית 100 פרטים. ידוע כי כל הברבורים מונוגמיים (בני הזוג נאמנים זה לזו), הנקבה מביאה לעולם רק צאצא אחד כל שנה, ונניח לצורכי שלמות הדוגמא כי האוכלוסייה יציבה, ללא הגירה וללא מחלות; שהברבורים בה חיים כולם 10 שנים והם מתרבים רק פעם אחת בחייהם בגיל חמש. עתה נשחק ונשנה את יחסי הזוויגים של 100 הפרטים באוכלוסייה ונחשב בכל דוגמא, מהו מספר הצאצאים שיוצר הזכר ומה מספר הצאצאים שיוצרת הנקבה. זאת משום שידוע שההצלחה האבולוציונית נמדדת על פי מספר הצאצאים "המוצלחים" אשר כל פרט מביא בדור הבא. לפי עקרון הברירה הטבעית, תכונה המגדילה את היכולת של הפרט להעמיד יותר צאצאים, להפרות יותר נקבות או הכושר להשיג יותר מזון – תלך ותיעשה נפוצה יותר באוכלוסייה עד שתהיה השלטת בה. לכן, מיצוי התיאוריה הדרווינסטית ושל עקרון הבררה הטבעית הוא מי שיש לו מספר צאצאים מוצלחים גדול יותר בדור הבא הוא המנצח במלחמת האבולוציה[8]. בטבלה 1 אנו מדגימים כיצד מחשבים את הכשירות הדרווינסטית של פרט באוכלוסיה הנוקט באסטרטגיה מסוימת (לדוגמה "בוחר" להיות זכר) . זוהי דוגמא היפותטית לכשירות (=התועלת) המתקבלת לכל זוויג ביחסים שונים באוכלוסיה בגודל נתון.
טבלה 1. דוגמאות לאופן החישוב של הכשירות הדרווניסטית של כל זוויג ביחסים שונים של שכיחויות זכרים ונקבות באוכלוסיה קבועה בת 100 פרטים.
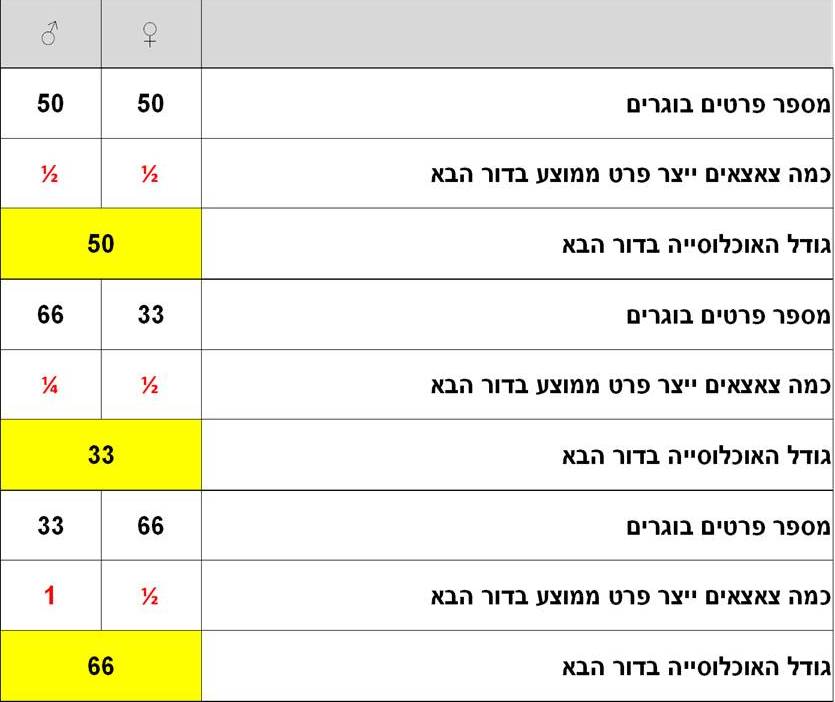
דוגמא 1: אוכלוסייה המורכבת מ- 99 נקבות וזכר בודד
במקרה זה, כל נקבה מטילה ביצה אחת ומכיוון שישנן 99 נקבות, אזי בדור הבא יהיו 99 צאצאים. יש לזכור שזכר יכול להפרות נקבות רבות. לפיכך החישוב הנכון צריך להתמקד בשאלה כמה צאצאים בממוצע מביא לעולם כל זוויג, ובחישוב כזה התועלת הדרוויניסטית שונה לחלוטין. כמה צאצאים מביאה כל נקבה? לכל נקבה צאצא בודד ולכן התועלת הדרוויניסטית שלה ב"ערכים של צאצאים" הוא רק חצי (0.5), שכן מבחינה גנטית הצאצא הוא חצי שלה וחצי של הזכר שהפרה אותה. כמה צאצאים מביא לעולם כל זכר במונחים של תועלת דרוויניסטית? ישנו רק זכר אחד; הוא הפרה את כל הנקבות; כלומר הכשירות הדרווינסטית שלו מחושבת ל- 99 צאצאים לחלק לשניים (כי מחצית מכל צאצא הוא גנטית שלו), דהיינו 49.5.
דוגמא 2: אוכלוסייה המורכבת מ- 99 זכרים ונקבה בודדת
במקרה זה, כמו במקרה הקודם, התועלת הדרוויניסטית של כל נקבה שווה לחצי. לעומת זאת, התועלת של זכר שווה רק לחצי של 1/99; מדוע ? מכיוון שהסיכוי של כל זכר להפרות את הנקבה הבודדת הוא רק 1/99. ומבחינה גנטית הצאצא נושא רק מחצית מהגנים שלו. לכן התועלת הצפויה לזכר היא רק 1/198.
לנוכח הדוגמאות הללו, ניתן להציג כעת את השאלה החשובה ביותר העומדת בבסיסה של האבולוציה: נאמר שעובר שזה עתה נוצר ו"רוצה" להוריש כמה שיותר מהגנים שלו לדור הבא, ויש לו אפשרות לבחור את הזוויג של עצמו – זכר או נקבה, מה היה מעדיף להיות: ילוד זכר או ילוד נקבה? התשובה בדוגמאות 1 ו-2 היא פשוטה וברורה: בדוגמא הראשונה כדאי להיות זכר, מכיוון שהתועלת הצפויה לזכר היא 49.5 בעוד שהתועלת של כל נקבה היא רק 0.5. לעומת זאת , בדוגמא 2 אם יש לעובר זכות בחירה, עדיף להיות נקבה שכן התועלת הדרוויניסטית שלה היא 0.5 (שים לב שתועלת הנקבה נשארת 0.5 בכל הדוגמאות) לעומת התועלת של זכר ממוצע שהפכה להיות זעומה (1/198 = 0.005~).
הקורא העירני ישאל את עצמו מה פירושה של "'זכות הבחירה" לכאורה של העובר. הרי התכונה להיות זכר או נקבה היא גנטית מעיקרה ונקבעת על פי הגנים וחוקי התורשה המנדלית ואיננה עניין של "בחירה חופשית". באמצעות דוגמא זו אנו רוצים להמחיש את התהליך האבולוציוני המתרחש בעובר: תוך כדי הפריית הביצית על ידי תא הזרע של הזכר, מתלכדים שני התאים ונוצר עובר. זוויג העובר נקבע על ידי חוקי התורשה של מנדל (ראו תיבה 2). באדם ובכל היונקים קיימים כרומוזומי מין מיוחדים הנקראים X ו-Y. לנקבה שני כרומוזומי XX ולזכר כרומוזום X יחיד וכרומוזום Y יחיד וביחד XY. מבנה גנטי זה יוצר בדיוק יחס צאצאים של 50:50. אך אם תתרחש מוטציה אשר תטה את יחס הזוויגים לטובת העובר[9] וכתוצאה מכך הכשרות הדרוויניסטית שלו תגדל – המוטציה תתפשט באוכלוסיה ובמשך האבולוציה תשתלט. זה מה שקורה כאשר יחס הזוויגים מוטה לאחד הצדדים- כאשר קיים רוב זכרים לעומת מעוט של נקבות וההפך – רוב של נקבות כנגד מיעוט של זכרים, כפי שראינו בדוגמאות מספר 1 ו-2. כלומר מבנה גנטי של אוכלוסייה בו המיניות נקבעת בכרומוזומי מין מיוחדים הוא המנגנון שהתפתח במשך האבולוציה כדי לקיים את שווי-המשקל האבולוציוני של יחס זוויגים שווה בו יש לשני המינים כשירות דרוויניסטית הגבוהה ביותר האפשרית. ההפרדה האקראית של כרומוזומי המין מביאה תמיד וליחס זוויגים מדויק של 50:50. אם תתרחש בעובר מוטציה אשר תטה את יחס הזוויגים לטובת אחד הזוויגים וכתוצאה מכך הכשירות הדרוויניסטית שלו תגדל, המוטציה תתפשט באוכלוסייה ובמשך הדורות תשתלט זה מה שקורה באותם המקרים בטבע שבהם יחס הזוויגים מוטה לאחד הצדדים – או שקיים רוב זכרים לעומת מיעוט של נקבות או להיפך, רוב של נקבות כנגד מיעוט של זכרים. אלה המקרים שהוצגו בדוגמאות 1 ו-2. בהמשך התהליך, הסלקציה היא תלוית תדירות, כאשר עם התקדמות הדורות עשוי להיווצר מצב שבו שוב לא יהיה עוד יתרון בגלל המוטציה שהתפשטה, ולמוטציה הפוכה כשתתרחש, יהיה יתרון.
| תיבה 2. מנגנון קביעת זוויג גנטי אקראי
בתיאור שהצגנו לעיל, קביעת טיפוס הזוויג היא תורשתית. כך הדבר באדם, בכל היונקים וגם בכל הציפורים. האדם נושא בגופו 44 כרומוזומים ועוד שני כרומוזומי מין. כאשר שני כרומוזומי המין הם מסוג XX הפרט הוא נקבה וכאשר הוא נושא כרומוזום X אחד ועוד כרומוזום Y אחד וביחד XY הפרט הוא זכר. מנגנון זה מכונה קביעת מין תורשתית מטיפוס XY. מחקרים הראו כי כרומוסום Y הוא הקובע זכריות וחסרונו (ולא כפל ה-X) קובע נקביות. במקרה של קביעת זוויג תורשתית נקבע מינו של העובר ברגע ההפריה של הביצית על ידי תא הזרע. כל תאי הגוף הם דיפלואידים כלומר נושאים בתוכם שני עותקים זהים כמעט של כל כרומוזום. לעומת זאת, תאי הביצית ותאי הזרע הם הפלואידים, כלומר מכילים רק מנה אחת של כל כרומוזום (הם נוצרו בחלוקת הפחתה, מיוזה). כל הביציות מכילות תמיד כרומוזום X אך תאי הזרע יכולים להכיל או כרומוסום מין X או Y. שכיחות תאי הזרע בעלי כרומוסום X זהה במדויק לשכיחות תאי הזרע בעלי כרומוסום Y, ועל כך מבוססת התורשה המנדלית של הזוויג. איזה תא זרע יפרה את הביצית? כזה הנושא Y או כזה הנושא X? זוהי שאלה של הסתברות ועל כן מנגנון קביעת הזוויג מתבסס על תורשה אקראית. אפשר לראות במנגנון זה מקבילה של 'מנגנון הגרלה של משלר ואומן" בתורת המשחקים אשר באמצעותו מתאפשר יצירת שני זוויגים המקבלים בשווי-משקל תשלומים זהים ( Shmida and Peleg, 1996 )
קביעת הזוויג באדם
|


דוגמא 3: אוכלוסייה המורכבת מ- 80 נקבות ו- 20 זכרים
מכיוון שיש רוב של 80 נקבות ורק 20 זכרים כל זכר יפרה בממוצע 4 נקבות לעומת הנקבה שהתועלת הדרוויניסטית שלה היא תמיד חצי. בדוגמא זו התועלת הדרוויניסטית של הזכר תהיה שווה ל 2= , כלומר התועלת שלו שווה ל- 2. גם בדוגמא זו יש יתרון להיוולד כזכר מכיוון שהתועלת הדרוויניסטית שלו גדולה מזו של הנקבה. כמו בדוגמא 1 לזכר תועלת יותר גדולה משל הנקבה, עם זאת אנו רואים שכאשר היחס בין הזכרים והנקבות מתקרב ל-50:50 ההפרש בין התועלות הדרוויניסטיות של הזכרים והנקבות מצטמצם.
מכיוון שהתועלת של הנקבה תמיד שווה לחצי באופן בלתי-תלוי ביחס בין הזכרים והנקבות באוכלוסייה, אין טעם לקחת את הדוגמא ההפוכה ( 20 נקבות ו-80 זכרים). במקום זאת נבדוק דוגמא נוספת שבה יחס הזכרים והנקבות קרוב יותר ל-50:50.
דוגמא 4: אוכלוסייה המורכבת מ- 40 נקבות ו-60 זכרים
מכיוון שיש רוב של 60 זכרים ורק 40 נקבות כל זכר יכול להפרות רק "שני-שליש"(2/3)נקבה, כלומר שהתועלת הדרוויניסטית שלו תהיה שווה לשליש(1/3). באוכלוסייה כזו יש יתרון לעובר הנולד כנקבה מכיוון שהתועלת הדרוויניסטית של הזכר היא שליש והיא קטנה מזו של נקבה השווה לחצי.
בדוגמאות 4,3,2,1 ראינו שיש יתרון לעובר אם הוא שייך לזוויג הנדיר באוכלוסייה, מכיוון שבזוויג זה התועלת הדרוויניסטית שלו תהיה גדולה יותר. ואכן הכללה זו נכונה: תמיד יש יתרון לזוויג בעל השכיחות הנדירה באוכלוסיה.
דוגמא 5: אוכלוסייה המורכבת מ- 50 נקבות ו-50 זכרים
כפי שכבר ראינו קודם, התועלת הדרוויניסטית של הנקבה שווה תמיד לחצי, אך ביחס זוויגים 50:50, התועלת הדרוויניסטית של הזכר תהיה גם היא שווה לחצי. מדוע? מכיוון שכל זכר באוכלוסייה זו מפרה בממוצע רק נקבה אחת, רק חצי גנום שייך לו, ולכן – התשלום הדרווינסטי שזכר זה מקבל בתועלת של מספר צאצאים הוא רק חצי. בכל האוכלוסייה נולדים בסך הכול חמישים צאצאים ממאה פרטים באוכלוסייה אשר 50 מהם זכרים ו-50 מהם נקבות. במצב זה התועלת הדרוויניסטית לכל זכר ונקבה שווה לחצי (0.5). יחס זה משקף את השוויון בין הזכרים לנקבות בדומה לתוצאות שספר ג'ון ארבותנוט בשנת 1710. בחברה כזו התועלת לעובר בבחירה, להיות נקבה או זכר שווה, וניתן להניח שלו יכלו היו מחציתם בוחרים להיות מזוויג אחד ומחציתם מהזוויג השני.כך נשמר יחס הזוויגים השווה בין זכרים לנקבות באוכלוסייה לאורך דורות.
מאידך נוכחנו כי במצב של דוגמא 5, נולדים באוכלוסייה רק 50 צאצאים בכל דור. מצב זה נחות בהרבה (כלומר יעיל למין הביולוגי הרבה פחות) מכל אוכלוסייה אחרת בה יהיה רוב לנקבות על פני הזכרים, שם ייוולדו לכלל האוכלוסייה הרבה יותר צאצאים (כמו למשל בדוגמא מס.1, נולדים לאוכלוסייה שבה זכר אחר ו-99 נקבות 99 צאצאים!) . לכאורה נראה כי מצב עם רוב של נקבות באוכלוסייה הוא מצב עדיף ומנצח והיעיל יותר לאוכלוסייה. אלא מאי? הפרטים באוכלוסייה "יריבו" כל הזמן על- "מי יהיה הזכר המאושר?", במצב זה תמיד תהיה סלקציה חזקה בין זוויגית ותוך אוכלוסייתית לטובת הגדלת מספר הזכרים עד ליחס זוויגים שווה. נוצר בפועל מצב של שווי-משקל פחות יעיל בו סך כמות הצאצאים שיוצרת האוכלוסייה הוא רק חצי מהיעילות המיטבית.
זהו למעשה היסוד העמוק של ה"קונפליקט של טובת הפרט לעומת טובת המין" אשר גילה דרווין והניאו-דרוויסטים הסבירו אותו בנוסחאות מתמטיות ובתהליך גנטי-אוכלוסייתי.
כאשר מדברים על מצב זה יעיל או לא יעיל, חשוב לייחס את היעילות לזהות ה"שחקן" הרלוונטי. יש הבדל גדול בין מצב יעיל לפרט או מצב יעיל לאוכלוסייה (לחברה, למין הביולוגי). הדוגמאות שהבאנו לעיל מדגימות יפה מתי נוצר הקונפליקט בין התועלת לפרט לבין התועלת למין (או לאוכלוסיה). הדוגמא של 99 נקבות וזכר אחד מבטאת את "טובת המין המיטבית" אך אין היא מצב יציב של שווי-משקל, כלומר הפרטים מטיפוס נקבי "יעדיפו" להפוך את הזוויג שלהם לזכר ולכן אין מצב זה יעיל להם. במקביל למצב בטבע, בכלכלה ובתורת המשחקים השיתופית מבחינים בין סבירות חברתית לסבירות אינדיבידואלית וכאשר נמצא פתרון המספק את שתי הדרישות הללו (כלומר שאין עליו "ערעור" : בכלכלה יוגדר המצב כ"פרטו אופטימל") – זהו המצב היציב או פתרון שווי-משקל (Peleg.and Sudhölter 2007.).
על יחס הזוויגים בצמחים: צמחים דו-זוויגיים (דו-מיניים), חד-זוויגיים (דו-ביתיים) וכאלה בעלי דגם זוויגיות גמיש
מרבית מיני הצמחים שלהם פרחים גדולים, הם בעלי פרחים דו-מיניים (הרמפרודיטים) ומכאן שכל פרט באוכלוסייה הוא דו-זוויגי (שמידע וכהן, 1983; שמידע, 1994, 1998; וולף וחבריו, 1995,; בוגין וחבריו, 1997). בצמחים בעלי פרחים זעירים, מואבקי חרקים או מואבקי רוח, שכיח ביותר טיפוס המיניות ההרמפרודיטי בו כל פרט באוכלוסייה נושא פרחים זכריים ופרחים נקביים (חד-ביתי). במקרים הללו אין שאלה של יחס הזוויגים שכן כל הפרטים באוכלוסייה הם דו-זוויגיים. ואולם, באחוז קטן מבין מיני הצמחים נמצא טיפוס זוויגיות דומה לזה השכיח בבעלי חיים. הפרטים בצמחים אלה הם חד- מיניים – זכרים או נקבות. טיפוס זוויגיות זה בצמחים נקרא "דו-ביתי" ובבעלי-חיים הוא נקרא "'חיה חד-זוויגית" (שמידע, 1998). האם גם במיני הצמחים הדו-ביתיים נשמר יחס הזוויגים 50:50? מסתבר שכן. מאז 1980 אנו מטיילים ברחבי ישראל וסופרים את יחס הזוויגים באוכלוסיות טבעיות של צמחים דו-ביתיים וברוב המינים מקבלים כי יחס הזוויגים שווה! כאשר לדוגמא ספרים מאה עצים או מטפסים לאורך שבילי החורש בגליל של ער אציל, אשחר ארץ-ישראלי, חרוב מצוי, קיסוסית קוצנית, שרביטן מצוי, אלה ארץ-ישראלית, טמוס מצוי, יחס הזוויגים נשמר בקפדנות בין 45 ל-55 לטובת זוויג כלשהו, וזאת תוך כדי טיול וספירה שטחית של פרטים בוגרים בלבד.



צמחים דו-מיניים (דו-זוויגיים)
מימין – כלנית מצויה. צילם: שיר ורד ©. בפרח אברי זכר (אבקנים) ואברי נקבה (עליים).
במרכז ומשמאל – אלון מצוי, צמח חד-ביתי בעל פרחי זכר ונקבה על אותו פרט. במרכז- תפרחות זכריות; צילמה: ערגה אלוני ©; משמאל – פרחים נקביים; צילם: ישי שמידוב ©
להגדלה – לחצו על התמונות




צמחים חד-זוויגיים (דו-ביתיים)
מימין לשמאל – אלת המסטיק זכר, פרחים אבקניים. צילם: גדי פולק ©; אלת המסטיק נקבה, פרחים. צילם: גדי פולק ©; שרביטן מצוי זכר, אצטרובלים. צילמה: ערגה אלוני ©; שרביטן מצוי נקבה, "פירות". צילמה: ערגה אלוני ©.
להגדלה – לחצו על התמונות
יוצא דופן הוא דבקון הזית. זהו צמח טפיל דו-ביתי אשר בו יחס הזוויגים מוטה לטובת הפרטים הנקביים. אך במקרה זה הסיבה ידועה: לדבקון מערכת כרומוזומלית מיוחדת היוצרת טבעת חישוקית בזמן המיוזה הגורמת להטיה של יחס הזוויגים (Calder and Bernhardt, 1983).


דבקון הזית – צמח דו-ביתי עם יחס זוויגים מוטה לטובת נקבות
מימין – צילם: גדי פולק ©; משמאל – צילם: אבי שמידע ©
גם במדבר ספרנו צמחים דו-ביתיים והופתענו לגלות כי יחס הזוויגים 50:50 נשמר ברובם: באוכלוסיות הטבעיות של תמר מצוי בקניוני מואב, שרביטן ריסני, סהרון משתלשל, מתנן שעיר, אוג קוצני ואלה אטלנטית בהר הנגב.


שרביטן ריסני. צילם: ישי שמידוב ©
מימין – צמח זכר בפריחה; משמאל – צמח נקבה עם אצטרובלים נקביים.
יוצא דופן הוא הערער האדום הגדל באלפיו בהרי אדום שבירדן. במין זה ניתן למצוא פרטים שהם "רק זכריים" פרטים שהם "רק נקביים" אך לרוב הפרטים "זוויג גמיש", כלומר הם בעלי יכולת לשנות את ההקצאה הזוויגית שלהם כפי שנמצא בערער קשקשי הגדל בערבות יוטה (Freemanet al 1980 , 1981). אנו משערים כי למיני הערער בקבוצת הערערים הקשקשיים אין כרומוזומי מין וקביעת טיפוס הזוויג שלהם מוכתבת מתנאי הסביבה, כמו שפרטנו במאמר בטבע הדברים ( שמידע וברנדייס, 2016). דגם דומה יש לרכפתן מדברי בו האוכלוסייה בנויה משלושה טיפוסי זוויג: "רק זכר", "רק נקבה" וטיפוס בעל "זוויג גמיש" (וולף וחבריו, 1995) רק מחקר גנומי-גנטי-אקולוגי עתידי, יאשש או יפריך את ההשערה על הקשר בין המבנה הגנטי של המינים הללו לבין השכיחות של טיפוסי הזוויג שלהם ויחס הזוויגים באוכלוסיות בטבע.


מימין – ערער אדום, ערער מקבוצת ה"קשקשיים". הזוויג גמיש. צילם: גדי פולק ©
משמאל – ערער גלעיני, ערער מקבוצת ה"מחטניים" – הזוויגים מופרדים. צילם: גדי פולק ©
אז מה למדנו מהדוגמאות?
במקרה של "משחק יחס הזוויגים" תמיד "כדאי" להיות בטיפוס הזוויג הנמצא בשכיחות הנדירה באוכלוסייה. זאת הבין פישר, המדען הגדול, כבר בשנת 1930, אם כי התיאור הפורמלי של "כלל פישר" על יחס הזוויגים נכתב רק בשנות השישים של המאה הקודמת ( Charnov 1978, 1982; Hamilton , 1967; Trivers and Hare, 1976).
לא תמיד טובת המין שווה לטובת הפרט. תהליך הברירה הטבעית הדרווינסטית פועל להגדלת תועלתו של כל פרט באוכלוסייה. באיזה מורפולוגיה והתנהגות הוא "יבחר". תהליך זה מוליך לסלקציה באוכלוסייה שבה שורדים ומתרבים הפרטים בעלי התכונות המקנות להם כשירות דרווינסטית טובה יותר שלפעמים גם תורמים למין כולו. דוגמת יחס הזוויגים, מתעלמת מתחרות בין מינית ומדגישה את הסיטואציה בה יש קונפליקט מובנה בין טובת המין לטובת הפרט. על ידי כך אנו נחשפים מחד למהותו של התהליך הדרווינסטי ומאידך לעקרונות בתורת-המשחקים ובכלכלה: השיקול באיזו אסטרטגיה לנקוט, (כלומר איזה טיפוס זוויג לבחור) תלוי במה יבחר השכן שלך. הבחירה של כל פרט היא אכן אופטימלית לו עצמו, אך כתוצאה ממנה יכולה האוכלוסייה ל"היתקע" למצבי שווי-משקל פחות יעילים. הרבה מההתנהגויות המכונות בבעלי-חיים ובצמחים בשם "אלטרואיסטיות" מדגימות למעשה את הקונפליקט אשר תארנו בדוגמת יחס הזוויגים.
ספרות:
בוגין ר וול ד ושמידע א 1997 יחס הזוויגים בעצי אלה ושכיחות העפצים בהם. אקולוגיה וסביבה 4: 9-3.
וולף ל אריאלי א וברנס ג ושמידע א 1995 טיפוסי מיניות והאבקה ברכפתן מדברי. אקולוגיה וסביבה 2: 78-71.
שמידע א וכהן ד 1983 זכר ונקבה ברא אותם – אבל למה? על המיניות בצמחים ובבעלי חיים. טבע וארץ כ"ו (1): 19-28.
שמידע א וברנדייס מ 2016 מדוע נכחדו הדינוזאורים: על אסטרטגיות של קביעת הזוויג וההשלכות שלהן על האבולוציה של צמחים וחיות. טבע הדברים 245: 24-14.
שמידע א 1994 טיפוסי מיניות בעצים ובשיחים. "אקולוגיה וסביבה" מס' 1:3 עמ' 186-177.
שמידע א 1996 מיניות האדם בראי האבולוציה: על ספרו של רובין בקר. גלילאו 19: 51-50.
שמידע א כהן ד ומוטרו ע 1996 אהבה צופנית – על ההבדלים בין פרחי זכר לבין פרחי נקבה בצמחים. ילקוט המכוורת 36: 20-19.
שמידע א 1998 בחירת בני-זוג בצמחים. כמעט אלפים 18: 13-10.
שמידע א וכהן ד 2016 מדוע קיימת רבייה מינית בטבע ? כתב יד.
————————————————————————————————-
Baker RR and Bellis MA 1995 Human Sperm Competition. Chapman & Nall, London.
Betzig L Muder M Band Turke P 1988 (eds.) Human Reproductive Behaviour, a Darwinian perspective. Cambridge U.P., Cambridge.
Calder M and Bernhardt P 1983 The Biology of Mistletoes. Academic Press, New-York.
Charnov EL 1982 The Theory of Sex Allocation. Princeton University Press, Princeton New Jersey.
Charnov EL 1978 Sex Ratio selection Eusocial hymenoptera. American Naturalist 112: 317-326.
Fisher RA 1930 The Genetical theory of Natural Selection. Dover.
Freeman DC Harper KT and Charnov EL 1980 Sex change in plants: old and new observations and new hypotheses. Oecologia 47: 222-232.
Freeman DC McArthur ED Harper KT and Blauer AC 1981 Influence of environment on the floral sex ratio of monoecious plants. Evo. 35: 194-197.
Haldane JBS 1932 The Cause of Evolution. Princeton University Press, Princeton.
Haldane JBS 1940 Science and Everyday Life. Macmillian, Ney York.
Hamilton WD 1967 Extraordinary sex ratios. Science, 156: 477-488.
Kendall MG and Plackett 1977 (eds.). Studies in the History and Probability, Vol.2, PP: 30-34, High Wycombe.[ Summary on John Abouthnot work]
Maynard-Smith J and Szathmary E 1995 The Major Transitions in Evolution. Freeman H H publisher, Oxford.
Peleg B and Sudhölter P 2007 ( Sec.edition). Introduction to the Theory of Cooperative Games, Springer, Berlin.
Ridley M (sec.Edition) 1996 Evolution. Blackwell Science, Abington, England.
Shmida A and Peleg B 1996 Strict and symmetric correlated equilibria are the distributions of the ESS's of biological conflicts with asymmetric roles. In: Understanding Strategic Interaction – Essays in Honor of Reinhard Selten, Albers W Guth W Moldovanu B and van Damme E (eds.) Springer -Verlag. pp. 149-170.
Trivers RL and Hare H 1976 Haplodiploidy and the evolution of social insects. Science, 191: 249-263.
הערות
[1] נכון שיש דוגמאות רבות לסטייה מיחס זה אך הן מהוות אחוז זעום מכלל המינים, במיוחד כששיקול טובת המין הביולוגי צופה לסטייה חזקה מיחס "חצי – חצי" , דהיינו שבאוכלוסיה יהיו מעט זכרים והרבה נקבות. ספרות ענפה נכתבה בנושא והקורא מוזמן לעיין ברשימת המקורות בסוף המאמר.
[2]שאלה נוספת ושונה ומעניינת היא מדוע ישנם רק שני זוויגים ? על כך קראו במאמרם של שמידע ופלג (Shmida and Peleg, 1995).
[3]הנני מודה לחברי פרופ' דוד וול על שהאיר את עינינו על מחקר זה ולימד אותנו את חשיבותו. ראה: Kendall & Plackett, 1977
[4] פרופ. רחל בן-אשר מעירה כי בגיל הנישואים יחס הזוויגים מתכנס ל-50:50 ועל כן לא נמצא באוכלוסיית בני-אדם בוגרת יותר נקבות מזכרים. תודתי נתונה לה על הערותיה המועילות לאורך כל המאמר.
[5] קיימים מחקרים רבים בעלי מספרים שונים ביחס לאחוז הנקבות והזכרים הנולדים, נתוני תמותה והיחס בדרגת העובר – וקצרה היריעה מלסכמם במאמר זה. ראה – Betzig et al., 1988.
[6]בשמותיהם הלועזיים: Ronald H. Fisher – מומלץ ספרו משנת 1930 (ראה ספרות); John B.S.Haldane – מומלץ ספרו משנת 1932.
[7]עולה השאלה המעניינת – מדוע ברוב היצורים יש רק שני טיפוסי זוויג. על "מתי קיימת סטייה אדפטיבית מיחס זוויגים שווה ?"- ראה Charnov 1982.
[8]לפי עקרון הבררה הטבעית , תכונה המגדילה את היכולת להעמיד יותר צאצאים – למשל כושר להפרות יותר נקבות, או כושר לייצר יותר צאצאים, או הכושר להשיג יותר מזון – תכונה זו תלך ותעשה נפוצה יותר באוכלוסיה עד שתהיה השלטת בה.
[9] לשם דיוק, צרכים היינו לכתוב "המוטציה תטה את יחס הזוויגים לטובת האסטרטגיה בה בחר העובר. ניתן לראות באפשרות הבחירה של העובר בין זוויג זכרי לנקבי משחק בו לכל שחקן (הוא העובר הבוחר בין שני זוויגים באמצעות מוטציה) שתי אסטרטגיות אפשריות. בדוגמא אנו מחשבים את התשלום שיקבל השחקן כאשר יבחר בכל אסטרטגיה. זוהי סלקציה תלוית תדירות שבה ככל שהמוטציה המתפשטת ונעשית שכיחה – יקטן יתרונה וייווצר יתרון למוטציה ההפוכה אם תתרחש.
תמונה בעמוד השער של כלנית

תמר מצוי (בר) בנחל זרד באדום. צילם: אבי שמידע ©
נספרו 200 פרטים, יחס הזוויגים הוא מחצית לזכרים ומחצית לנקבות ונמצא מתאים בדיוק לתיאוריה.
תודה ליצחק אילון (דישל) ממושב כפר יחזקאל, אשר לימד אותנו לספור את הנקבות והזכרים של התמרים בגנים התלויים של קניוני ירדן. ראו: רווק, ש. ושמידע, א. 2013: אל קניוני מואב ואדום. הוצאת "טבע הדברים", עמודים 32-33, 122-123
=======================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: שמידע א קריינר ע וקריינר ח 2016 מדוע יחס הזוויגים השכיח בטבע ברוב האורגניזמים הוא 50:50?, כת-עת "כלנית" מספר 3.
https://www.kalanit.org.il/?p=6888&preview=true









