

תמר קיסר החוג לביולוגיה וסביבה, אוניברסיטת חיפה – אורנים tkeasar
עדי שדה מו"פ דרום, חוות הבשור adisadeh.a.s@gmail.com
אבי שמידע מחלקה לאבולוציה, סיסטמטיקה והתנהגות והמרכז לרציונליות, האוניברסיטה העברית, ירושלים avi.shmida
תקציר: פרחים שונים, הגדלים על אותו הצמח, מכילים לעתים קרובות כמויות שונות מאוד של צוף. הבדלים אלה עשויים לנבוע מפעילות שיחור מזון של חרקים מאביקים בפרחי הצמח. לחלופין, הם עשויים לשקף את השפעתה של הברירה הטבעית, המקנה יתרון לשונות גבוהה בייצור צוף בין פרחים. שונות זו צפויה לקצר את רצף הביקורים של מאביקים 'שונאי-סיכון' בצמח יחיד, לגרום להם לעבור לצמח שכן ובכך להפחית את הסיכון להאבקה עצמית. במחקר זה בדקנו את תרומתם של המאביקים ושל הצמחים עצמם לשונות ביבול הצוף בשיח רוזמרין רפואי Rosmarinus officinalis (משפחת השפתניים), המואבק בעיקר על ידי דבורים. מדדנו את קצב ייצור הצוף, יבול הצוף (כמות הצוף בפרחים) ואת ביקורי החרקים בשלושה צמחים מאוכלוסייה אחת במהלך 17 ימים בעונת פריחה אחת. מצאנו שונות גבוהה בקצבי ייצור הצוף (מקדם השונות=1.48), שהייתה גדולה יותר לאחר ימים גשומים. השונות ביבול הצוף הייתה גבוהה עוד יותר (CV=2.16), היא פחתה עם הטמפרטורה והתגברה עם הזמן שעבר מאז הגשם האחרון. קצב הביקורים של דבורים בפרחים פחת ככל שההבדלים ביבול הצוף בין פרחים גדלו, עלה עם העלייה במספר הפרחים הפתוחים על השיח, אבל לא הושפע מהשונות בקצב הייצור של הצוף בפרחים. דגימה חוזרת של פרחים מסומנים, שרוקנו מצוף, לא הראתה מתאם מובהק בין יבולי הצוף בפרחים לבין קצבי ייצור הצוף לאחר מכן. מתוצאות אלה אנו מסיקים, ששונות גבוהה ביבול הצוף בין פרחים באותו צמח מפחיתה את מספר ביקורי הדבורים בשיח לפני שיעברו לשיח אחר. אולם לצמחים אין בקרה מלאה על שונות זו, אלא היא מווסתת בעיקר על ידי שיחור המזון של המאביקים. הבקרה החלקית של מאביקים על השונות בכמויות הצוף צפויה להפחית את היתרון האבולוציוני לצמח מיצירת הבדלים בגמול הצוף בין פרחים שונים על אותו צמח.
=================
מבוא
צמחים מואבקי-חרקים, המבוקרים על ידי פרטים רבים של מאביקים, נהנים מתועלת רבייתית הכוללת ייבוא וייצוא של אבקה. החרקים מביאים אבקה ומפרים את הפרח, ובו בזמן לוקחים ממנו אבקה להפרייה של פרחים אחרים. מבחינים בין האבקה זרה, שבה מקור האבקה הוא בפרט אחר של אותו מין ביולוגי בעל הרכב גנטי שונה, לבין האבקה עצמית או האבקת שכנים (גיטונוגמיה). בהאבקה עצמית והאבקת שכנים, פרח מואבק על ידי אבקה של עצמו או של פרחים שכנים על אותו צמח, שלהם הרכב גנטי דומה. שני המקרים מובילים להאבקה ולהפריה עצמית, השקולים לנישואי אחים במשפחה ועלולים לגרום ליצירת צאצאים פגומים. בחלק ממיני הצמחים קיימים מנגנוני אי-התאם עצמי, המונעים הפריה במקרים של האבקה עצמית או האבקת שכנים. בין אם קיים התאם עצמי ובין אם לא, האבקה עצמית והאבקת שכנים מביאות לפגיעה בעשיית הזרעים, המשקפת את ההצלחה הרבייתית הנקבית של הצמח. האבקת שכנים עלולה לפגוע גם בהצלחה הרבייתית הזכרית כתוצאה מייצוא מופחת של אבקה לצמחים זרים. לכן הועלתה הסברה, שתכונות צמחיות המפחיתות את שכיחות האבקת השכנים מקנה לצמח יתרון אבולוציוני (de Jong et al., 1993; Reusch, 2001; Barrett, 2003). כשיש שפע של מאביקים, המצב האופטימלי עבור הצמחים הוא לקבל ביקורים מפרטים רבים של חרקים, שכל אחד מהם יבקר בכל צמח פעמים מעטות ככל האפשר אבל יעבור בין צמחים רבים בני אותו המין (Klinkhamer et al., 1994).
לעתים קרובות, מאביקים מבקרים רק בחלק קטן מהפרחים העומדים לרשותם על צמח מסוים, ואז הם עוזבים אותו ועוברים לפרט אחר (Klinkhamer and de Jong, 1990; Ohashi and Yahara, 2001). ניסויים במעבדה ובשדה הראו, שנחיתה על פרחים חסרי גמול מזון, או בעלי גמול נמוך, גורמת לדבורים לעבור בין כתמי מזון שונים (Kadmon and Shmida, 1992; Keasar et al., 2002). יתר על כן, דבורים לעיתים קרובות מראות 'שנאת-סיכון' בעת שיחור למזון, כלומר הן מעדיפות מקורות מזון בעלי שונות נמוכה על פני מקורות בעלי תוחלת גמול שווה אבל שונות גבוהה יותר (Kacelnik and Bateson, 1996; Shafir, 2000). תצפיות אלה מעלות את ההיפותזה, ששונות בייצור הצוף בין פרחים על אותו הצמח עשויה להפחית את שכיחות האבקת השכנים, ולכן יהיה לה יתרון אבולוציוני (Rathcke, 1992; Pappers et al., 1999; Biernaskie et al., 2002). על פי היפותזה זו, שונות בייצור הצוף בין פרחים שונים תגביר את השונות ביבול הצוף על הצמח. הגברת השונות עשויה להפחית את מספר הביקורים הרצופים של המאביקים בכל פרט של הצמח ובכך להקטין את שיעור האבקת השכנים.
מבחינים בין יבול צוף, שהוא כמות הצוף (הנמדדת במיקרוליטר) אותה מסוגל המאביק לשאוב מכל פרח בו הוא מבקר, לבין ייצור צוף (הנמדד במיקרוליטר ליחידת זמן) שהוא קצב הפרשת הצוף על ידי צופני הפרח לאחר שרוקן מהצוף. במחקרים קודמים, שנערכו במיני צמחים אחדים, נמדדו הבדלים גדולים בקצב ייצור הצוף (בהיעדר צריכה על ידי חרקים) בין פרחים שעל פני אותו הצמח (Feinsinger, 1983; Zimmerman and Pyke, 1986; Herrera and Soriguer, 1983; Marden, 1984, Boose, 1997). קיימת גם שונות וכתמיות מרחבית בתוך הצמח ביבול הצוף של פרחים (Shmida and Kadmon, 1991). יבול הצוף בפרחים (הנמדד בפרחים הזמינים לביקורי חרקים) מושפע מקצב הייצור על ידי הפרח ומקצב הצריכה שלו על ידי מאביקים. דבורים משתמשות באסטרטגיה של חיפוש מוגבל-אזור כשהן אוספות צוף, כלומר עפות מרחקים קצרים לאחר ביקור בפרח עשיר בגמול ומרחקים ארוכים יותר לאחר ביקור בפרחים בעלי גמול נמוך (Keasar et al., 2006, Burns and Thomson, 2006). דגם תנועה זה מביא לניצול כתמי של הצוף, ויוצר כתמים עשירים וכתמים דלים בצוף, גם אם לא הייתה קיימת כתמיות ביבול הצוף מלכתחילה (Motro and Shmida, 1995). לכן הקשר בין השונות בייצור הצוף לבין השונות ביבול הצוף אינו מובן מאליו. השונות ביבול הצוף בתוך צמח עשויה לנבוע מדגם ייצור הצוף על ידי הצמח, מדגם הצריכה שלו על ידי מאביקים, או משילוב של שני הגורמים. לכן, דרושה מדידה משולבת של קצב ייצור הצוף, יבול צוף בפרחים ופעילות המאביקים כדי לבחון את הסוגיות הבאות: האם מקורה של השונות ביבול הצוף, המוצע כגמול למאביקים, הוא בצמח או בחרקים? האם שונות זו מקצרת את רצפי הביקורים של מאביקים בפרחים של צמח מסוים ועשויה להפחית בכך את שכיחותה של האבקת השכנים?
במחקר זה מדדנו את קצב ייצור הצוף, את יבול הצוף ואת תדירות ביקורי המאביקים בשיחים של רוזמרין רפואי (.Rosmarinus officinalis L) ממשפחת השפתניים (תמונה 1). הרוזמרין הוא שיח מואבק דבורים בעל תפוצה מערב ים-תיכונית המגודל כצמח נוי בישראל. בישראל הוא פורח בסוף החורף ובאביב, וכל שיח מייצר בו-זמנית מאות פרחים. האבקנים בפרחים מבשילים תחילה, ורק לאחר מכן הופכת הצלקת זמינה להפרייה (פרחים פרוטנדריים). הפרחים הם בעלי התאם עצמי, כלומר אין בהם מנגנון גנטי המונע הפרייה עצמית. לכן סובל הצמח מפגיעה בכשירות כתוצאה מהאבקה והפריית שכנים (Hidalgo and Ubera, 2001). בכל יום תצפית רשמנו את מספר הפרחים הפתוחים לשיח, מפני שגודל הצמח עשוי להשפיע על קצב ייצור הצוף (Pleasants and Chaplin, 1983), על יבול הצוף ולכן גם על קצב ביקורי המאביקים (Brody and Mitchell, 1997; Goulson et al., 1998). בנוסף, אספנו נתונים על אירועי גשם לפני ימי התצפית ועל הטמפרטורה במהלך התצפיות מפני שמשתנים אלה עשויים להשפיע על קצב ייצור הצוף וקצב ביקורי המאביקים (Wyatt et al., 1992; Carroll et al., 2001; Leiss and Klinkhamer, 2005).
שאלות המחקר העיקריות היו:
- מהי השונות בקצב ייצור הצוף בפרחים ומהי השונות ביבול הצוף אותו פוגשים המאביקים בפרחים שונים של אותו שיח?
- האם השונות ביבול הצוף משפיעה על קצב הביקורים של מאביקים?

תמונה 1. רוזמרין רפואי. צילום: אבי שמידע ©
חומרים ושיטות
איסוף הנתונים
מדדנו את קצב ייצור הצוף ואת ביקורי המאביקים בשלושה שיחי רוזמרין רפואי בקיבוץ חצור. השיחים לא קיבלו השקייה לאורך תקופת המחקר. הרוזמרין היה המין הדומיננטי שפרח במקום בין אוקטובר למרץ. אורך ורוחב הכותרת היו 1.37±0.18 ס"מ (ממוצע±סטיית תקן) ו 0.50±0.07 ס"מ בהתאמה, ואורך צינור הפרח היה 1.04±0.25 ס"מ (n=50). נתוני הצוף וביקורי המאביקים נאספו במשך 17 ימים בתקופה אוקטובר-דצמבר 2002, בשעות 14:00-8:00 בתחילת כל יום תצפית (בשעה 8) מדדנו את יבול הצוף ב 10 פרחים מכל שיח באמצעות פיפטות בנפח 1μl. הפרחים שנבחרו למדגם היו 5 זוגות של שכנים קרובים ממקומות שונים על השיח. חישבנו את המתאם ביבול הצוף בין בני הזוג, ובין פרחים מזוגות שונים, כמדד לכתמיות פיזור הצוף בתוך השיח. לאחר מכן כיסינו את הפרחים שרוקנו מצוף בשקיות מבד כילה (Wyatt et al., 1992) ובדקנו את נפחי הצוף שהצטברו בהם כעבור שלוש שעות (11:00), שש שעות (14:00) ו 24 שעות (8:00). בכל פעם דגמנו 10 פרחים (4-3 מכל שיח) ומדדנו את נפחי הצוף בהם, המייצגים את ייצור הצוף בפרחים לאורך 6,3 ו 24 שעות. חישבנו את קצב ייצור הצוף לשעה על ידי חלוקת נפח הצוף שנמדד בזמן שעבר ממועד הכיסוי של הפרחים. בדגימות שנפחן עלה על 1/3 מיקרוליטר מדדנו את ריכוז הסוכר בצוף בעזרת רפרקטומטר ידני מתוצרת Bellingham-Stanley. מכשיר זה אינו מאפשר מדידה אמינה של דגימות שנפחן נמוך מ 1/3 מיקרוליטר. ריכוז הסוכר נבדק בדגימות יבול צוף וייצור צוף כאחד. רשמנו את שעת הבדיקה כדי לאפיין קשר אפשרי בין זמן המדידה לבין ריכוז הצוף.
ספרנו את ביקורי החרקים במדגם של כ 100 פרחים על כל שיח במשך 10 דקות באותם שיחים ששימשו למדידות צוף. התצפיות נערכו בין 12-10 בבוקר, שהיו שעות השיא של פעילות המאביקים. בכל יום ערכנו תצפית אחת בכל שיח, ובסה"כ 51 תצפיות. מיינו את החרקים המבקרים לקבוצות הפונקציונליות הבאות: דבורי דבש, דבורים גדולות (יותר מדבורי דבש), דבורים קטנות (יותר מדבורי דבש), זבובים, פרפרים וחיפושיות.
ניתוח הנתונים
השתמשנו בניתוח שונות כדי לבחון את השפעת משך הכיסוי של הפרחים על קצבי ייצור הצוף לשעה. התייחסנו למשתנה 'שיח' כאל גורם אקראי בניתוח השונות, מפני שקיימת תלות בין דגימות חוזרות של ייצור צוף מאותם הצמחים. בחנו את השפעת תאריך הדגימה, השיח הנדגם וזוג הפרחים בתוך השיח על קצב ייצור הצוף באמצעות מודל GLM. התייחסנו לתאריך הדגימה (17 ערכים אפשריים) כאל גורם קבוע, ואל מספר השיח (3 ערכים אפשריים) כאל גורם אקראי. מספר זוג הפרחים (5 ערכים אפשריים, שקול למיקום על השיח) הוגדר כגורם מקונן בתוך שיח ובתאריך דגימה נתון. ערכנו מבחני פירסון כדי לזהות קורלציות בקצב ייצור הצוף בין פרחים בני-זוג.
יבול הצוף נמדד בפרחים שלא כוסו בכילות, ולכן בדקנו כיצד הושפע מתאריך הדגימה, מספר השיח ומספר זוג הפרחים בלבד. גם כאן השתמשנו במודל GLM שבו התאריך (17 ערכים אפשריים) הוגדר כגורם קבוע, מספר השיח (3 ערכים) הוגדר כגורם אקראי ואילו מספר זוג הפרחים (5 ערכים) הוגדרו כגורם מקונן בתוך תאריך ושיח.
השתמשנו בניתוחי רגרסיה לינארית כדי לבחון את השפעתם של משתני הסביבה הבאים על יבול וייצור הצוף: עצמת הגשם האחרון (מ"מ גשם באירוע הגשם האחרון לפני התצפית), מספר הימים שעברו מאז אירוע הגשם האחרון, טמפרטורת מינימום ומקסימום בזמן התצפית. חישבנו את יבול וייצור הצוף הממוצעים, עם סטיות התקן שלהם, עבור כל אחת מ 51 התצפיות (17 ימים × 3 שיחים) של ביקורי חרקים. חישבנו רגרסיות של מספר ביקורי החרקים בכל תצפית כנגד (1) יבול הצוף הממוצע (2) סטיית התקן של יבול הצוף (3) מספר הפרחים הפתוחים על השיח. קיימת תלות בין נקודות הנתונים ברגרסיות הללו, מפני שמקורן במדידות חוזרות על אותם שלושה שיחים. לכן בדקנו את השפעת הצמח (גורם אקראי) על מספר ביקורי החרקים בכל תצפית. השתמשנו בגרפים של רגרסיות חלקיות כדי להראות את השפעת מספר הפרח והשונות ביבול הצוף על קצב ביקורי החרקים, אחרי שהבאנו בחשבון את השפעת השיח. השתמשנו בטרנספורמציה לוגריתמית כדי לנרמל את נתוני יבול הצוף וייצור הצוף טרם הניתוח הסטטיסטי. הפעלנו טרנספורמציית שורש ריבועי על נתוני ביקורי המאביקים. הניתוחים הסטטיסטיים בוצעו בתכנת SPSS גרסה 15.0.
תוצאות
ייצור צוף
נפחי הצוף הממוצע (±סטיית תקן) שיוצרו במשך 3, 6 ו 24 שעות היו 0.17±0.24μl, 0.18±0.26μl ו 0.43±0.60μl, בהתאמה. בהתאם לכך, קצב ייצור הצוף לשעה היה הגבוה ביותר בפרחים שכוסו בכילות למשך 3 שעות (בין 8 ל 11 בבוקר), והנמוך ביותר בפרחים שכוסו למשך 24 שעות (בין 8 בבוקר ל 8 למחרת) (איור 1). השפעת משך הכיסוי על קצב ייצור הצוף לשעה הייתה מובהקת ביותר (F2, 335=45.90, P<0.001). קצבי ייצור הצוף לשעה נבדלו באופן מובהק בין זוגות פרחים, אך לא בין שיחים ובין תאריכי דגימה. עם זאת, נמצאה אינטראקציה מובהקת בין המשתנים 'תאריך הדגימה' ו 'שיח' (GLM: F15,366=1.38, P=0.16 עבור תאריך הדגימה; F2,366=1.37, P=0.26 עבור השיח, F30,366=3.20 P<0.0001 עבור האינטראקציה תאריך×שיח, F4,366=3.18, P=0.01 עבור זוג פרחים; משך הכיסוי הוגדר כ covariate). לא נמצאה אינטראקציה מובהקת בין משך הכיסוי לבין השיח (F2,366=0.53, P=0.59) ותאריך הדגימה ( F15,366=1.01, P=0.44).
36% מהפרחים שכוסו למשך שלוש שעות, 39.4% מהפרחים שכוסו למשך שש שעות ו 31.2% מהפרחים שכוסו למשך 24 שעות לא ייצרו צוף בכמויות מדידות במשך כל תקופת הכיסוי.
 איור 1. הקצב הממוצע של ייצור הצוף בפרחים שכוסו למניעת ביקורי מאביקים למשך 6,3 ו-24 שעות. אורך הקופסא מייצג את טווח הרבעון. החציון מצויין בקו עבה בתוך כל קופסא והעמודות מציינות את סטיית התקן. כל ממוצע מייצג מדידות של 170 פרחים. עמודות המצוינות באותיות דומות אינן שונות באופן מובהק על פי מבחן Tukey Post-Hoc.
איור 1. הקצב הממוצע של ייצור הצוף בפרחים שכוסו למניעת ביקורי מאביקים למשך 6,3 ו-24 שעות. אורך הקופסא מייצג את טווח הרבעון. החציון מצויין בקו עבה בתוך כל קופסא והעמודות מציינות את סטיית התקן. כל ממוצע מייצג מדידות של 170 פרחים. עמודות המצוינות באותיות דומות אינן שונות באופן מובהק על פי מבחן Tukey Post-Hoc.
בהמשך התייחסנו רק לנתוני ייצור הצוף אשר התקבלו מפרחים שכוסו לאורך שלוש שעות. התעמקנו בנתונים אלה משתי סיבות: ראשית, ייתכן שדגם ייצור הצוף המוצג באיור 1 נובע מקצבי הפרשה גבוהים יותר בין השעות 11-8 בבוקר מאשר בהמשך היום. מכיוון שפעילות המאביקים הייתה בשיאה בשעות אלה, סברנו שקצבי ייצור הצוף בשעות הבוקר עשויים להשפיע על מירב ביקורי המאביקים, ובכך תתאפשר בדיקת ההשפעות היחסיות של הצמחים לעומת המאביקים על השונות ביבול הצוף. שנית, ייתכן שהצטברות צוף בצינור הפרח מעכבת הפרשה נוספת, ולכן קצב הייצור פוחת כשזמן הכיסוי של הפרחים מתארך. הצטברות כזו לא צפויה תחת תנאים טבעיים, מפני שהפרחים זוכים לביקורים תכופים של מאביקים (כמדווח להלן). לכן נתונים מפרחים שכוסו למשך זמן ארוך (6 או 24 שעות) עלולים לתת הערכת חסר של קצב הייצור ברוזמרין רפואי.
מצאנו שונות גבוהה (מקדם השונות CV = 1.48, n=170 פרחים) בקצבי ייצור הצוף לשעה (החישובים מבוססים על פרחים שכוסו למשך שלוש שעות). ריכוז הסוכר הממוצע (w/w) בפרחים היה 70.6±8.5% (מקדם השונות = 0.12, n=65 פרחים). השונות בריכוז הצוף הייתה נמוכה באופן מובהק מאשר השונות בנפח (F117,64=10.04, P<0.001, מבחן Lewontin 1966 לשוויון בין מקדמי השונות). נמצאה קורלציה מובהקת בקצבי ייצור הצוף בין פרחים בני-זוג על פני אותו הצמח (n=85 זוגות פרחים, מקדם פירסון = 0.39, P<0.001). קצבי הייצור הושפעו באופן מובהק מכמות הגשם האחרון לפני הדיגום. מספר הימים מאז הגשם האחרון וטמפרטורות המינימום והמקסימום ביום הדיגום לא השפיעו על קצבי ייצור הצוף (טבלה 1).
טבלה 1. רגרסיה מרובה לבדיקת התלות של קצב ייצור הצוף בפרחי רוזמרין רפואי. החלק העליון מציג את התוצאות הכלליות של מודל הרגרסיה והחלק התחתון את ההשפעה של כל אחד ממשתני הסביבה: כמות המשקעים בגשם האחרון, מספר הימים מאז הגשם האחרון, טמפרטורת המינימום וטמפרטורת המקסימום ביום הבדיקה על קצב ייצור הצוף.
| P | F | Mean square | df | Sum of squares |
Model |
|
| 0.034 | 2.692 | 0.020 | 4 | 0.078 |
Regression |
|
| 0.007 | 125 | 0.910 |
Residual |
|||
| 129 | 0.989 |
Total |
| המשתנה |
t |
Coefficient |
P |
|
| כמות הגשם האחרון | 3.070 | 0.327 | 0.003 | |
| מס הימים לאחר הגשם | 0.643 | 0.082 | 0.521 | |
| טמפרטורת מינימום | 0.100 | 0.014 | 0.921 | |
| טמפרטורת מקסימום | 0.272 | 0.045 | 0.786 |
יבול הצוף לפרח היה 0.32±0.69μl (ממוצע±סטיית תקן, n=510 פרחים, CV=2.16), כלומר הייתה שונות גבוהה בין פרחים ביבול הצוף. 56% מהפרחים לא הכילו צוף כלל, בעוד ש- 1.4% מהדגימות הכילו יותר מ 3μl צוף, מה שמעיד על התפלגות לא-סימטרית של יבולי הצוף בין פרחים. מקדם השונות של יבולי הצוף היה גבוה באופן מובהק מאשר מקדם השונות של קצבי ייצור הצוף לשעה (F75, 117=2.14, P<0.001, מבחן Lewontin (1966) לשוויון מקדמי השונות). נמצא קשר חיובי מובהק בין יבולי הצוף של פרחים בני זוג, בדומה לממצא עבור קצבי ייצור צוף (n=255 זוגות פרחים, מקדם פירסון = 0.42, P<0.001). היו הבדלים מובהקים ביבולי הצוף בין תאריכי דגימה, אבל לא בין צמחים או זוגות פרחים (GLM: F16,267=22.68, P<0.001 עבור תאריך דגימה; F2,407=1.24, P=0.29 עבור שיח; F12,267=0.69, P=0.76 עבור אינטראקציה תאריך דגימה × שיח; F222,267=01.05, P=0.35 עבור זוג פרחים). לא נמצא מתאם בין יבולי הצוף לבין קצב ייצור הצוף שנמדדו באותם הפרחים (מתאמי פירסון: r=0.05, P=0.49; r=-0.06, P=0.45; r=0.07, P=0.34 עבור פרחים שכוסו במשך שלוש, שש ו 24 שעות בהתאמה). יבולי הצוף פחתו עם טמפרטורת האוויר המקסימלית, ועלו עם הזמן מאז הגשם האחרון, אבל לא הושפעו מכמות הגשם באירוע האחרון ומטמפרטורות האוויר המינימליות (טבלה 2).
טבלה 2. רגרסיה מרובה לבדיקת התלות של יבול הצוף בפרחי רוזמרין רפואי החלק העליון מציג את התוצאות הכלליות של מודל הרגרסיה והחלק התחתון את ההשפעה של כל אחד ממשתני הסביבה: כמות המשקעים בגשם האחרון, מספר הימים מאז הגשם האחרון, טמפרטורת המינימום וטמפרטורת המקסימום ביום הבדיקה על יבול הצוף.
| P | F | Mean square | df | Sum of squares |
Model |
|
0.049 |
2.452 | 4.238 | 4 |
16.951 |
Regression |
| 1.729 | 147 |
254.103 |
Residual |
||
| 151 |
271.054 |
Total |
| המשתנה | t | Coefficient | P |
| כמות הגשם האחרון | -1.166 | -0.105 | 0.246 |
| מס הימים לאחר הגשם | 2.230 | 0.264 | 0.027 |
| טמפרטורת מינימום | 1.898 | 0.237 | 0.060 |
| טמפרטורת מקסימום | -2.719 | -0.429 | 0.008 |

איור 2. רגרסיה חלקית לבדיקת ההשפעה של מספר הפרחים הפורחים לשיח על מספר הביקורים של מאביקים ל 100 פרחים במשך 10 דקות. מודל הרגרסיה כלל גם את מספר הצמח, הממוצע וסטיית התקן של יבול הצוף כמשתנים בלתי תלויים. הקשר בין מספר הפרחים למספר הביקורים היה מעריכי (אקספוננציאלי), והמשתנים עברו טרנספורמציית לוג לקבלת פונקציה של קו ישר. שיפוע הגרף הוא המעריך של הפונקציה המקורית, המקשרת בין מספר פרחים למספר ביקורים (de Jong & Klinkhamer, 2005). מספר התצפיות N=51, ונוסחת ישר הרגרסיה היא Y=0.663x+36.17, r2=0.09, P<0.001.
ביקורי מאביקים
צפינו ב 63.4±8.1 (ממוצע±סטיית תקן) ביקורי מאביקים (נחיתה על הפרח) ל 100 פרחים לאורך 10 דקות (n=51 תצפיות). נתון זה שקול ל 3.8 ביקורים לפרח לשעה בממוצע. לא רשמנו הגעה של מאביקים ללא נחיתה על הפרחים, ואת משכי הביקורים. דבורי דבש היו המין המבקר הנפוץ ביותר (57.1% מכל הביקורים), 36.3% מהביקורים נערכו על ידי דבורים קטנות, ויתר קבוצות המאביקים ביקרו בפרחים בתדירויות נמוכות. מספר ביקורי החרקים עלה באופן מובהק עם מספר הפרחים על השיח (איור 2, t50=2.30, P=0.03), אך לא נבדלו באופן מובהק בין שיחים (t50=-0.64, P=0.53). מספר ביקורי החרקים ירד עם העלייה במקדם השונות של יבול הצוף (איור 3, t50=-2.34, P=0.03) ולא הושפעו מהבדלים בין השיחים (t50=0.34, P=0.73). יבולי הצוף הממוצעים לא השפיעו על מספר ביקורי החרקים (t50=-1.75, P=0.09). ברגרסיה זו, כמו בניתוחים הקודמים, לא מצאנו השפעה של השיח על מספר ביקורי החרקים (t50=0.47, p=0.64).
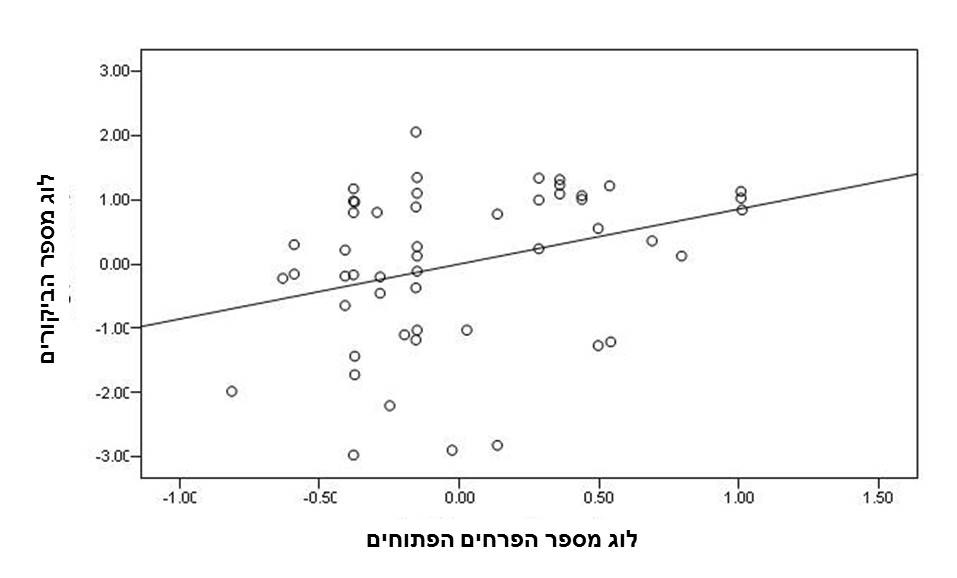
איור 3. רגרסיה חלקית לבדיקת ההשפעה של השונות (מיוצגת על ידי סטיית התקן) ביבול הצוף בפרחים על מספר הביקורים של מאביקים ל 100 פרחים במשך 10 דקות. מודל הרגרסיה המלא כלל גם את מספר הצמח, יבול הצוף הממוצע ומספר הפרחים על השיח כמשתנים בלתי תלויים. N=51 תצפיות, ונוסחת ישר הרגרסיה היא: Y=-3.98x+72.48, r2=0.08, P=0.047.
דיון
במחקרנו אספנו נתונים אודות השונות בייצור צוף ויבול צוף בפרחים, במשולב עם נתונים על ביקורי מאביקים, בסדרת תצפיות בשדה. התצפיות שערכנו ברוזמרין רפואי מציגות שונות גבוהה בקצבי ייצור צוף בין פרחים של אותו הצמח ושונות גדולה אף יותר ביבול הצוף של הפרחים, כפי שמעיד מקדם השונות הגבוה יותר. נמצא קשר הפוך בין קצב ביקורי המאביקים לבין השונות ביבול הצוף, אך לא נמצא קשר בין קצב ביקורי המאביקים לשונות בקצב ייצור הצוף. מתוצאות אלה ניתן להסיק, שקצב ייצור הצוף של הצמחים לא היה המקור היחיד לשונות ביבול הצוף במחקרנו. לכן, אנו משערים ששיחור המזון של החרקים המאביקים העצים את השונות בתוך השיחים, מעבר לשונות בייצור הצוף שמקורה בצמחים. שונות גבוהה זו, בתורה, הקטינה את מספר ביקורי המאביקים. במילים אחרות, השונות ביבול הצוף עשויה להיות גם סיבה וגם תוצאה של דגם ביקורי החרקים בפרחים. עם זאת, כשליש מהפרחים לא ייצרו צוף כלל לאורך הזמן שבו היו מכוסים. פרחים ריקים אלה הם בבירור חלק מתרומתם של הצמחים לשונות ביבולי הצוף, מפני שאין למאביקים אפשרות לשנות את יבול הצוף בפרחים שאינם מכילים גמול כלל.
הסבר אפשרי לתוצאות הוא שהבדלים בקצבי ייצור הצוף בין פרחים יוצרים שונות התחלתית ביבול הצוף. זו מפחיתה את מספר ביקורי החרקים באותו שיח ומקנה יתרון אדפטיבי לצמח על ידי צמצום סיכוני ההפריה העצמית כתוצאה מהאבקת שכנים. לכן צפויה במקרה זה ברירה טבעית להגברת השונות בייצור צוף על ידי פרחים שונים באותו צמח. בהמשך, ביקורי מאביקים עשויים להגדיל עוד יותר את השונות ביבול הצוף, על ידי יצירת כתמים מקומיים של פרחים ריקים מצוף. במקרה כזה, בו המאביקים מגיבים בעיקר לשונות ביבול הצוף שהם עצמם יצרו בזמן השיחור למזון, לא צפויה לפעול ברירה טבעית לטובת שונות בייצור צוף על ידי פרחים שונים באותו הצמח.
חוסר אחידות בקצב ייצור צוף בין פרחים עשוי לשקף גם הבדלים בין פרחים בשלב זוויגי (Carlson and Harms, 2006), בגיל (Pleasants, 1983; Zimmerman and Pyke, 1986) או בתנאי בית הגידול. למשל, מכיוון שהפריחה ברוזמרין רפואי מתקדמת מתחתית התפרחת אל ראשה, ייתכנו הבדלים בגיל ובשלב הזוויגי בין פרחים במקומות שונים על גבי התפרחת, וכתוצאה מכך גם בקצבי ייצור צוף.
מצאנו מתאם חיובי בקצב ייצור צוף וביבול צוף בין פרחים שכנים בצמח, אך לא נמצא מתאם בין קצב ייצור הצוף לבין יבול הצוף בפרח בודד. במילים אחרות, הפרחים שהכילו את נפחי הצוף הגדולים ביותר כשהיו חשופים לביקורי מאביקים, לא ייצרו צוף בקצב הגבוה ביותר לאחר שכוסו בכילות. הסבר אפשרי לממצא זה הוא, שקצב ייצור הצוף משתנה במהלך חיי הפרח, ושמדידה של ייצור ויבול הצוף בנקודות זמן שונות משקפת את השינויים הללו. בתמיכה לכך, נמצא במספר מיני צמחים שגיל הפרח משפיע על קצב ייצור הצוף (Southwick and Southwick, 1983; Galetto et al., 1994). לחלופין ניתן לשער, שחלק מהשונות ביבול הצוף נובעת מפעילות מאביקים, בעוד שהשונות בייצור הצוף נמצאת תחת בקרת הצמח בלבד. כדי להבחין בין שתי הפרשנויות יש לבצע מדידות חוזרות של קצבי ייצור הצוף לאורך חייהם של פרחים מסומנים.
מחקרנו מאשש תוצאות של עבודות קודמות, שמצאו השפעות של משתני סביבה על תכונות צוף (Wyatt et al., 1992; Carroll et al., 2001; Leiss and Klinkhamer, 2005). מעניין לציין שקצב ייצור הצוף ויבול הצוף ברוזמרין הושפעו ממשתנים שונים. הגשם היה המנבא הטוב ביותר של קצבי ייצור צוף, בעוד שטמפרטורה ניבאה בצורה הטובה ביותר את יבולי הצוף. טמפרטורות גבוהות מאפשרות פעילות מאביקים מוגברת, מה שעשוי להגדיל את צריכת הצוף ולהפחית את היבול. השפעה אפשרית נוספת של טמפרטורות גבוהות היא נידוף מוגבר של מים מהצוף, וכתוצאה מכך הקטנת הנפחים הנמדדים. אולם, נידוף מוגבר צפוי להביא גם לעליה בריכוזי הצוף, ואילו אנחנו לא מצאנו הבדלים בריכוז הצוף בשעות שונות. לכן אנו משערים שלנידוף מים הייתה השפעה מוגבלת בלבד על נפחי הצוף שמדדנו. בניגוד לתוצאות של מחקר קודם (Pleasants and Chaplin, 1983), לא מצאנו השפעה של מספר הפרחים על השיח על קצב יצור הצוף ועל יבולו. מצד שני, מספר הפרחים הפתוחים השפיע על משיכת מאביקים, כפי שנמצא בעבודות קודמות במיני צמחים אחרים (Robertson and McNair, 1995; Goulson et al., 1998).
מצאנו קשר שלילי בין השונות ביבול הצוף לבין ביקורי חרקים בשיחי המחקר, אך לא בין השונות בייצור הצוף לבין תדירות ביקורי החרקים. ייתכן שהשונות בייצור הצוף לא מספיקה כדי להשפיע על ביקורי חרקים, וששונות זו ממוסכת על ידי השונות הגבוהה יותר ביבול הצוף. השונות בייצור הצוף מושפעת מהצמח בלבד, בעוד שהשונות ביבול מושפעת מיצור הצוף וגם מפעילות המאביקים. המשמעות היא, שלשונות שנוצרת כתוצאה מפעילות החרקים יש השפעה חשובה על התנהגותם, ושחשיבות השונות הנוצרת על ידי הצמח מצטמצמת. אם כך, תרומת המאביקים לשונות ביבול הצוף צפויה להקטין את היתרון הסלקטיבי של שונות הנוצרת על ידי הצמח כאסטרטגיה המפחיתה האבקת שכנים.
תודות
עבודת השדה נתמכה על ידי הקק"ל. ניתוח הנתונים והכתיבה נתמכו על ידי קבוצת המחקר על אבולוציה ותורת המשחקים במכון ללימודים מתקדמים, האוניברסיטה העברית. תום דה-יונג העיר על כתב היד המקורי באנגלית.
מאמר זה פורסם במקור באנגלית:
Keasar, T., Sadeh, A. and Shmida, A. 2008. Variability in nectar production and yield, and their relation to pollinator visits, in a Mediterranean shrub. Arthropod-Plant Interactions 2: 117-123
———————————–
ספרות:
Barrett SCH 2003 Mating strategies in flowering plants: the outcrossing–selfing paradigm andbeyond. Philosophical Transactions of the Royal Society of London Series B 358: 100–991.
Biernaskie JM Cartar RV and Hurly TA 2002 Risk-averse inflorescence departure in hummingbirds and bumble bees: could plants benefit from variable nectar volumes? Oikos 98: 98–104.
Boose DL 1997 Sources of variation in floral nectar production rate in Epilobium canum (Onagraceae): implications for natural selection. Oecologia 110: 493–500.
Brody AK and Mitchell RJ 1997 Effects of experimental manipulation of inflorescence size on pollination and pre-dispersal seed predation in the hummingbird-pollinated plant Ipomopsis aggregata. Oecologia 110: 86–93.
Burns JG and Thomson JD 2006 A test of spatial memory and movement patterns of bumblebees at multiple spatial and temporal scales. Behavioral Ecology 17: 48–55.
Carlson JE and Harms KE 2006 The evolution of gender-biased nectar production in hermaphroditic plants. Botanical Review 72: 179–205.
Carroll AB Pallardy SG and Galen C 2001 Drought stress, plant water status, and floral trait expression in fireweed, Epilobium angustifolium (Onagraceae). American Journal of Botany 88: 438–446.
Castellanos MC Wilson P and Thomson JD 2002 Dynamic nectar replenishment in flowers of Penstemon (Scrophulariaceae). American Journal of Botany 89: 111–118.
Galetto L Bernardello M and Juliani HR 1994 Characteristics of secretion of nectar in Pyrostegia venusta (Ker-Gawl.) Miers (Bignoniaceae). New Phytologist 127: 465–471.
de Jong TJ and Klinkhamer PGL 2005 Evolutionary Ecology of Plant Reproductive Strategies. Cambridge University Press, Cambridge, New York.
de Jong TJ Waser NM and Klinkhamer PGL 1993 Geitonogamy: the neglected side of selfing. Trends in Ecology and Evolution 8: 321–325.
Feinsinger P 1983 Variable nectar secretion in a Heliconia species pollinated by hermit hummingbirds. Biotropica 15: 48–52.
Goulson D Stout JC Hawson SA and Allen JA 1998 Floral display size in comfey, Symphytum officinale L (Boraginaceae): relationships with visitation by three bumblebee species and subsequent seed set. Oecologia 113: 502–508.
Herrera CM and Soriguer RC 1983 Intra- and inter-floral heterogeneity of nectar production in Helleborus foetidus L. (Ranunculaceae). Botanical Journal of the Linnean Society 86: 253–260.
Hidalgo PJ and Ubera JL 2001 Inbreeding depression in Rosmarinus officinalis L. International Journal of Developmental Biology 45(S1): S43–S44.
Kacelnik A and Bateson M 1996 Risky theories—the effects of variance on foraging decisions. American Zoologist 36: 402–434.
Kadmon R and Shmida A 1992 Departure rules used by bees foraging for nectar: a field test. Evolutionary Ecology 6: 142–151.
Keasar T Shmida A and Motro U 1996 Innate movement rules in foraging bees: flight distances are affected by recent rewards and are correlated with choice of flower type. Behavioral Ecology and Sociobiology 39: 381–388.
Keasar T Rashkovich E Cohen D and Shmida A 2002 Bees in two-armed bandit situations: foraging choices and possible decision mechanisms. Behavioral Ecology 13: 757–765.
Klinkhamer PGL and de Jong TJ 1990 Effects of plant size, plant density and sex differential nectar reward on pollinator visitation in the protandrous Echium vulgare (Boraginaceae). Oikos 57: 399–405.
Klinkhamer PGL de Jong TJ and Metz JAJ 1994 Why plants can be too attractive – a discussion of measures to estimate male fitness. Journal of Ecology 82: 191–194.
Leiss KA and Klinkhamer PGL 2005 Genotype by environment interactions in the nectar production of Echium vulgare. Functional Ecology 19: 454–459.
Lewontin RC 1966 On the measurement of relative variability. Systematic Zoology 15: 141–142.
Marden JH 1984 Intrapopulation variation in nectar secretion in Impatiens capensis. Oecologia 63: 418–422.
Motro U and Shmida A 1995 Near-Far search: an evolutionarily stable foraging strategy. Journal of Theoretical Biology 173: 15–22.
Ohashi K and Yahara T 2001 Behavioural responses of pollinators to variation in floral display size and their influences on the evolution of floral traits. In: Chittka L and Thomson J D (eds) Cognitive Ecology of Pollination. Cambridge University Press, Cambridge, New York, pp 274–296.
Pappers SM de Jong TJ Klinkhamer PG L and Meelis E 1999 Effects of nectar content on the number of bumblebee approaches and the length of visitation sequences in Echium vulgare (Boraginaceae). Oikos 87: 580–586.
Pleasants JM 1983 Nectar production patterns in Ipomopsis aggregata(Polemoniaceae). American Journal of Botany 70: 1468–1475.
Pleasants JM and Chaplin SJ 1983 Nectar production rates of Ascelapias quadrifolia: causes and consequences of individual variation. Oecologia 59: 232–238.
Rathcke BJ 1992 Nectar distributions, pollinator behavior, and plant reproductive success. In: Hunter M D Ogushi T Price P W (eds) Effects of Resource Distribution on Animal–plant Interactions. Academic Press, NY. pp 113–138.
Reusch TBH 2001 Fitness consequences of geitonogamous selfing in a clonal marine angiosperm (Zostera marina). Journal of Evolutionary Biology 14: 129–138.
Robertson AW and MacNair MR 1995 The effects of floral display size on pollinator service to individuals flowers of Myosotis and Mimulus. Oikos 72: 106–114.
Shafir S 2000 Risk-sensitive foraging: the effect of relative variability. Oikos 89: 663–669.
Shmida A and Kadmon R 1991 Within-plant patchiness in nectar standing crop in Anchusa strigosa. Plant Ecology 94: 95–99.
Southwick AK and Southwick EE 1983 Aging effect on nectar production in two clones of Asclepias syriaca. Oecologia 56: 121–125.
Wyatt R Broyles SB and Derda GS 1992 Environmental influences on nectar production in milkweeds (Ascelapias syriaca and A. exaltata). American Journal of Botany 79: 636–642.
Zimmerman M and Pyke GH 1986 Reproduction in Polemonium: patterns and implications of floral nectar production and standing crops. American Journal of Botany 73: 1405–1415.
===================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: קיסר ת שדה ע ושמידע א 2016 שונות ביבול וייצור צוף, והשפעתם על ביקורי מאביקים ברוזמרין רפואי, שיח ים-תיכוני, כתב-עת "כלנית" 3.
https://www.kalanit.org.il/rosmarinus-pollination/









