

גידי נאמן, החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים gneeman@gmail.com
כאשר עץ גדול של אורן ירושלים נשרף, הוא מת ואינו מתחדש באופן וגטטיבי, בניגוד למרבית עצי החורש. במקום בו היה עץ אורן גדול שנשרף כליל, נותר עיגול אפר גדול. לאפר pH גבוה בסיסי המונע, כמעט לחלוטין, נביטה של צמחים, בחורף הראשון לאחר שריפה. למרות זאת, מספר קטן של זרעי אורן, מצליחים לנבוט באותו מקום בו היה העץ השרוף. בשל העדר תחרות בין- ותוך-מינית, והזנה המינרלית משופרת אותה מספק האפר, הזריעים המועטים, שורדים וצומחים מהר יותר מזריעי האורנים בשאר השטח. בהמשך עתידים הנבטים הללו להחליף את העץ השרוף שבאתרו הם נבטו. כתוצאה מכך, המבנה המרחבי של היער המתחדש לאחר שריפה, יהיה דומה לזה של היער שנשרף. בניגוד לצפוי, בדיקה גנטית שללה את ההשערה שהעצים המתחדשים באתר של עץ אורן שרוף הם רק צאצאיו. לכן, ההרכב והמבנה הגנטי המרחבי של היער המתחדש לאחר שריפה דומה, אבל אינו זהה לזה של היער שנשרף.
מבוא
שריפות משפיעות על האקולוגיה והאבולוציה של צמחים במקומות רבים בעולם. הקיץ הארוך, החם והיבש מאוד במקומות בעלי אקלים ים-תיכוני בעולם (אגן הים-התיכון, קליפורניה, אוסטרליה, דרום אפריקה וצ'ילה) גורם לריבוי שריפות שחלקן טבעיות מברקים (לא בישראל) ומרביתן מעשה ידי אדם (Biswell 1974 Naveh 1994, Trabaud 1994, Keeley et al. 2011). במשך 2000-1980 נשרפו למעלה מ 107 דונם בדרום אירופה (European Commission, 2001). בישראל נשרפו ב 1991-1980 כ 136,000 דונם (קליאוט וקידר 1992). בשריפה הגדולה בדצמבר 2010 בכרמל, נשרפו כ 25,000 דונם של יערות אורן וחורשים (פרבולוצקי ושות 2011) (איור 1).

איור 1. שריפות יער בכרמל, צילום: שי לוי ©
שריפות משמידות את מרבית הביומסה הצמחית מעל לפני הקרקע, הן גורמות לשינויים במבנה הפיזי של הקרקע, לירידה בכמות החומר האורגני ובתכולת המינרלים בקרקע (Mataix-Solera et al. 2011). שריפות משפיעות על בנק הזרעים בקרקע (Ne’enan & Izhaki 1999) ועל נביטת הזרעים (Trabaud and Oustric 1989, Moreno and Oechel 1991, Thanos et al. 1992).
הצמחייה, באזורים בעלי אקלים ים-תיכוני, מותאמת לקיץ יבש נטול משקעים ועמידה בפני שריפות (Keeley 1986, Malanson 1987). העמידות ויכולת ההתחדשות של הצמחים היא תוצאה של התאמות ייחודיות המאפשרות השתקמות של אוכלוסיות הצמחים שנכחו טרום השריפה (Enright et al. 2014). מרבית המינים שורדים, ומתחדשים ווגטטיבית לאחר שריפה, מניצנים בבסיס הגזע, או על ענפים (Clarke et al. 2013, Pausas & Keeley 2017). ההתחדשות מתרחשת מספר שבועות לאחר השריפה, גם בקיץ לפני גשמי החורף. מינים אחרים, כמו מרבית מיני האורן Pinus sp. ומיני הלוטם Cistus sp., מתים לאחר שריפה, ואינם מתחדשים ווגטטיבית. האוכלוסיות של מינים אלה, מתחדשות רק באמצעות נביטת זרעים מבנק הזרעים בקרקע, או השמור בצמרת העץ (Lamont et al. 1991, Pausas et al. 2004). נביטת הזרעים מתרחשת לאחר גשמי החורף הראשונים. הנביטה מכוונת ומווסתת על-ידי גורמים הקשורים לשריפה, כמו חשיפת הזרעים היבשים לטמפרטורה גבוהה ועשן (Keeley & Fotheringham 1997). לאחר הנביטה, יש תחרות חזקה, בין הנבטים הגדלים בצפיפות, על מרחב, מים ואור (Ne’eman 1994). להכוונת הנביטה, לצמיחה וההתבססות של הצמחים לאחר השריפה יש השפעה מכרעת על נישת ההתחדשות של המינים השונים, על הדמוגרפיה של האוכלוסיות, ועל הפיזור המרחבי של הפרטים באוכלוסיות לאחר השריפה (Ne’eman 1997).
פיזור מרחבי של מינים ופרטים בסקלה מקומית, נחקר ונמצא בעל חשיבות אקולוגית (Shmida and Whittaker 1981, Auerbach and Shmida 1987). התחדשות צמחים לאחר שריפה, תוארה, במרבית המקרים, מנקודת מבט של השינויים החלים עם חלוף הזמן מאז השריפה האחרונה. השינויים בהרכב ומגוון המינים ובתצורת הצומח, זאת בקנה מידה גדול של חברת הצמחים והנוף (Naveh 1989, Trabaud 1994). ברור שלפיזור המרחבי של הצמחים לפני השריפה, יש השפעה על דגם ההתאוששות של הצמחים אחריה, אולם, רק מחקרים ספורים תיעדו עובדה זו (Davis et al. 1989, Rice 1993) או ניסו להבין את המכניזמים והמשמעות האקולוגית של התופעה (Moreno & Oechel 1991, Ne’eman 1997). הדגם המרחבי של הצמחים לאחר שריפה מושפעת בו זמנית ממספר גורמים:
1) הפיזור המרחבי של הצמחים לפני השריפה המשפיע על הדגם וההרכב של בנק הזרעים בקרקע.
2) בצמחים בעלי בנק זרעים השמור באצטרובלים אפילים (סרטוניים) בצמרות העצים, המפזרים זרעים בעיקר לאחר שריפה, הפיזור המרחבי של העצים הבוגרים טרם השריפה קובע את הדגם של הפצת הזרעים לאחריה.
3) הפיזור המרחבי של הביומסה הצמחית לפני השריפה, משפיע על עוצמת השריפה, ועל מידת החימום של הקרקע. לחימום הקרקע השפעה על התמותה של זרעים הטמונים בה ועל הנביטה של זרעים השורדים את החימום. לכן , הוא משפיע על הדגם המרחבי של חברת הצמחים לאחר שריפה ועל צפיפות הפרטים באוכלוסייה.
4) הפיזור המרחבי של הביומסה הצמחית טרום השריפה קובע את כמות האפר. לאפר תגובה בסיסית מאוד המשפיעה באופן דיפרנציאלי על הנביטה של מינים שונים (Henig-Sever et al. 1995).
5) השונות המרחבית בעוצמת השריפה ובכמות האפר, יוצרת מגוון של תנאים מקומיים לאחר השריפה. שונות זו משפיעה על מידת התחדשות של צמחים ויוצרת דגם של מוזאיקה נופית לאחר השריפה, כפי שתועד לאחר השריפה הגדולה ב 1988 בילוסטון ארה"ב (Turner & Romme 1994).
אורן ירושלים Pinus halepensis Miller הוא מרכיב חשוב של יערות ברום נמוך באגן הים-התיכון (Barbéro et al. 1998, Quézel 2000) שעיקר תפוצתו היא מערב ים-תיכונית. בישראל קיימת אוכלוסייה טבעית, מבודדת ונפרדת מבחינה גנטית, שהוגדרה כזן המזרחי של אורן ירושלים (Korol et al. 2002). אורן ירושלים הוא עץ דליק מאוד המת בזמן שריפת צמרות, אולם האוכלוסייה מתחדשת היטב אחריה מנביטת זרעים בלבד (Ne’eman and Trabaud 2000, Saracino & Leone 1993, Pausas 2015 ). מקורם של מרבית הנבטים הוא מזרעים האגורים באצטרובלים אפילים serotinous (איור 2), המפזרים זרעים בעיקר לאחר התייבשותם בזמן שריפה (Lamont et al. 1991, Nathan et al. 1999). אחוז האצטרובלים האפילים גבוה יותר בעצים צעירים מאשר מבוגרים ובאוכלוסיות שהתחדשו באופן טבעי לאחר שרפות, יחסית לאוכלוסיות שהתבססו בשטחים מופרים ללא שריפה (Goubitz et al. 2004). תכונות אלה מקטינות את סכנת ההיכחדות של אוכלוסיות צעירות (juvenility risk) במקרה של שריפה חוזרת, לפני שהספיקו לייצר בנק זרעים בגודל מספיק להתחדשות (Keeley et al. 2004). זרעים מאצטרובלים אפילים מותאמים טוב יותר לנביטה לאחר שריפה מזרעים באצטרובלים רגילים, שכן הם עמידים יותר לחימום ולריכוז אפר גבוה בקרקע (Goubitz et al. 2003).


איור 2. איצטרובלים רגילים פתוחים ואפילים סגורים ואצטרובל אפיל שנפתח לאחר שריפה, צילום: גידי נאמן ©


במקרים רבים נמצא מתאם בין מיקום הצמרות של עצים בוגרים ומקום הנביטה של צאצאיהם (Horn 1975, Frelich et al. 1993), דבר המגדיל את הסיכוי שאתר המתאים לגידול צמח בוגר יישאר מאוכלס על-ידי אותו מין גם בעתיד. לאחר שריפה, צמחים המתחדשים וגטטיבית ממשיכים לחיות באותו מקום, אבל מה קורה עם צמחים המתים בשריפה ומתחדשים אחריה מזרעים בלבד? מה הסיכויים של עץ, כמו אורן ירושלים, לנבוט ולהתבסס באתר טוב המסוגל להצמיח עץ בוגר? על מנת לענות על שאלה זו בדקנו את הפיזור המרחבי של הנביטה של זרעי אורן ירושלים .Pinus halepensis Mill. ואת שיעור ההישרדות והצמיחה של הזריעים לאחר שריפה, יחסית למיקום הצמרות של העצים הגדולים שנשרפו ומתו.
בשלב שני, ביקשנו לבדוק את ההשפעה של השריפה על ההרכב הגנטי, השונות הגנטית והמבנה הגנטי המרחבי של אוכלוסייה אורן ירושלים המתחדשת לאחריה. האם המבנה הגנטי המרחבי של אוכלוסיית האורנים המתחדשת לאחר שריפה דומה לזה של הדור הקודם שהוכחד בשריפה? במיוחד ביקשנו לבדוק, האם הזריעים של האורנים הנמצאים מתחת לצמרות שרופות של עצים גדולים הם גם הצאצאים שלהם?
שיטות
אתר המחקר
המחקר נערך ביערות טבעיים לא נטועים בפארק הכרמל. האקלים הוא ים-תיכוני טיפוסי עם ממוצע שנתי של כ- 700 מ"מ גשם שמרביתם יורדים בחודשים דצמבר-פברואר. המסלע בכל אתרי המחקר הוא שכבות של נארי אשר בתנאי האקלים הנתונים מייצר קרקע רנדזינה עשירה בגיר. מרבית יערות האורן הטבעיים בכרמל הם תוצאה של התפשטות טבעית, שמקורה במספר קטן יחסית של צמחי אם שפלשו לשטחי חקלאות ומרעה. שטחים שננטשו בעיקר בתחילת המאה העשרים, בדומה למצב במערב אגן הים-התיכון (Barbéro et al.1998). יערות אלה הם רב גילים בצפיפות של כ 20 עצי גדולים לדונם. מתחתם תת-יער של חורש צפוף הכולל בעיקר עצים של אלון מצוי וחרוב מצוי, שיחים של אלת מסטיק וקידה שעירה. בשטחים הפתוחים יותר, גדלים בעיקר בני שיח של לוטם מרווני ולוטם שעיר, סירה קוצנית, גאופיטים ומינים עשבוניים רב- וחד-שנתיים רבים (איור 3).

איור 3. אורנים טבעיים גדולים בכרמל עם חורש מתחתם צילום: גידי נאמן ©

שיטות המחקר
במחקר זה בדקנו את הדגם המרחבי של נביטת הזרעים של אורן ירושלים לאחר שריפה, צמיחתם והתבססותם יחסית לצמרות השרופות של עצים גדולים שמתו בשריפה. בכל עץ גדול שרוף ערכנו ארבעה חתכים מאונכים של ריבועים בשטח של מ"ר כל אחד. גבול הצמרות השרופות היה במרחק ממוצע של כארבעה מ' מהגזע. לכן הגדרנו כל ריבוע שהיה במרחק של עד ארבעה מטרים כ 'קרוב', כלומר מתחת לצמרת השרופה. כל ריבוע במרחק גדול מארבעה מ' מהגזע כ 'רחוק', כלומר מחוץ לצמרת השרופה. בכל ריבוע נספרו כל הזריעים, ונמדדו הגובה והקוטר של כל אחד מהם. לכל זריע חושב 'מדד הביומסה' (הקוטר X הגובה בריבוע) אשר שיקף באופן מובהק את משקל הזריעים (R2=0.94, P<0.05). חישבנו את הממוצעים עבור כל ריבוע, ועבור כל הריבועים שגדלו קרוב או רחוק מהעץ הסמוך. לאחר מכן, ערכנו השוואה בין מדדי הביומסה של זריעים שגדלו קרוב לבין אלה שגדלו רחוק מהגזע המת. השוואה כזו עשינו בארבעה אתרים: שני אתרים שהיו שנתיים וחמש שנים לאחר שריפה סמוך לחי-בר.11 אתר של שנים לאחר שריפה ברכס מתלה ואתר 20 שנים לאחר שריפה ברכס אצבע. כולם יערות אורן טבעיים שלא טופלו, וניתן היה לזהות בהם את מיקום הגזע של העצים הגדולים שנשרפו.
על מנת לבדוק את השפעת השריפה על המבנה הגנטי, השונות הגנטית והפיזור המרחבי של זריעי אורן ירושלים לאחר שריפה, השווינו בין אוכלוסייה צעירה שהתחדשה באופן טבעי לאחר שריפה ברכס לובים, לאוכלוסייה סמוכה שלא נשרפה. בדקנו, גם, האם ניתן היה להבחין במבנה גנטי מרחבי מסוים בקרב אוכלוסיית הזריעים המתחדשת, ומה הם הסיכויים שהזריעים הגדולים ביותר הם גם צאצאים של העצים השרופים שמתחתם הם נבטו. במקור ביקשנו להשוות את ההרכב הגנטי של הנבטים לזה של העצים השרופים שמעליהם. מכיוון שלא הצלחנו בכך, בדקנו האם הזריעים הגדולים מתחת לעץ מסוים הם בעלי אם משותפת. לשם כך, דגמנו את הנבטים, שצמחו מתחת לצמרות השרופות של 20 עצים גדולים, שנה וחצי לאחר שנשרפו. מתחת לכל עץ דגמנו את עשרת הנבטים הגדולים ביותר שהיו במרחק של שני מ' לפחות מהגזע השרוף. זאת מתוך הנחה שרבים הסיכויים שצמח אחד מתוך הקבוצה יחליף את העץ הגדול שמת בשריפה. מכל נבט מוצה ה DNA ותשעה אתרים של מיקרוסטליטיים מוכרים בעלי שונות גדולה חוזקו ב PCR. מכיוון שלא הצלחנו לבודד DNA מהעצים השרופים השווינו את ההרכב הגנטי של הנבטים בכל קבוצה ובין הקבוצות תוך שימוש באתר ITPH4516 (קטע DNA) שנמצאו בו 15 אללים שונים. השוואה זו לא אפשרה לבדוק את ההשערה שלנו האם העץ השרוף היה האם של הנבטים, אבל אפשר לנו לבדוק האם לכל הנבטים בקבוצה היה לפחות אלל אחד משותף. אם היה כזה, זו עדות לאם משותפת ואם לא זה אומר שהנבטים בקבוצה מקורם בזרעים של לפחות שתי אימהות נפרדות. פרטים נוספים על שיטות העבודה ניתן למצוא במאמרים המדעיים המקורים (ראו בתחילת המאמר).
תוצאות
דמוגרפיה


איור 4. התחדשות אורן ירושלים מנבטים לאחר שריפה קרוב לגזע שרוף (מימין) ורחוק ממנו (משמאל) צילום: אנה גרשברג וגידי נאמן ©

טבלה 1. מדד הביומסה והצפיפות (מספר זריעים למ"ר) הממוצעת וסטיית התקן של זריעי אורן שצמחו קרוב (4מ'>) או רחוק (4מ'<) מהגזעים השרופים. n הוא מספר העצים ו P המובהקות של ניתוח שונויות ANOVA להשוואה בין הממוצעים.
| קרוב | קרוב | רחוק | רחוק | ||||
| מדד ביומסה | שנים | ממוצע | ס.ת. | ממוצע | ס.ת. | P | n |
| חי-בר א' | 2 | 0.001 | 0.0006 | 0.001 | 0.0001 | 0.07 | 41 |
| חי-בר ב' | 5 | 0.1 | 0.11 | 0.06 | 0.1 | 0.03 | 62 |
| מיתלה | 11 | 7.72 | 6.5 | 0.45 | 0.53 | 0.0001 | 82 |
| אצבע | 20 | 16.96 | 11.2 | 1.94 | 1.7 | 0.0001 | 58 |
| צפיפות | |||||||
| חי-בר א' | 2 | 8.23 | 7.16 | 12.87 | 8.32 | 0.0001 | 60 |
| חי-בר ב' | 5 | 4.04 | 2.15 | 3.12 | 2.3 | 0.002 | 76 |
| מיתלה | 11 | 0.67 | 0.34 | 0.86 | 0.47 | 0.02 | 82 |
| אצבע | 20 | 0.93 | 0.53 | 0.97 | 0.55 | 0.69 | 53 |
בניתוח שונויות דו כיווני נמצאה השפעה מובהקת של האתר (שנים מאז שריפה) (F3=663.1, P<0.0001) ושל המיקום (קרוב או רחוק מהגזע( (F1=140, P<0.0001) על מדד הביומסה של הזריעים. גם האינטראקציה בין המיקום והאתר הייתה מובהקת (F2=26.5, P<0.0001), כלומר השפעת המיקום הייתה שונה באתרים שונים. שנתיים לאחר השריפה לא היה הבדל בביומסה של הזריעים הקרובים לרחוקים, אבל מאוחר יותר הזריעים שגדלו רחוק היו גדולים בהרבה מאלו שגדלו קרוב לגזע השרוף (טבלה 1).
בניתוח שונויות דו-כיווני נמצאה השפעה מובהקת של האתר (שנים מאז שריפה) (F3=153.7, P<0.0001) אבל לא של המיקום (קרוב או רחוק מהגזע( (F1=2.6, P>0.1) על צפיפות הזריעים. גם האינטראקציה בין המיקום והאתר הייתה מובהקת (F2=7.7, P<0.0001), כלומר השפעת המיקום הייתה שונה באתרים שונים. שנתיים לאחר השריפה צפיפות הנבטים קרוב לגזע השרוף הייתה קטנה בהרבה מזו של הנבטים שצמחו רחוק מהגזע. ההבדל היה קטן יותר לאחר חמש ו 11 שנים, ולאחר 20 שנים הבדל זה נמוג (טבלה 2).
למעט שנתיים אחרי שריפה, למרחק מהגזע הייתה השפעה מובהקת על הביומסה של הזריעים, שהייתה גבוהה במיוחד 11 ו 20 שנים לאחר שריפה (איור 5).

איור 5. רגרסיה לינארית בין מדד הביומסה של זריעי אורן ירושלים לבין המרחק מהגזע השרוף חמש שנים (A, n=690), 11 שנים (B, n=590) ו 20 (C, n=590) שנה לאחר שריפה. n= מספר הריבועים.
הרכב ומבנה גנטי
על פי בדיקת הרכב הגנטי של האתר ITPH4516 בקטע ד.נ.א, שנמצאו בו 15 אללים שונים, מרבית קבוצות הנבטים (11 מתוך 20) שגדלו כל אחת מתחת לעץ אורן גדול שרוף לא היו בעלי אם משותפת (העץ שמתחתיו נבטו), והיו להם לפחות שתי אימהות שונות. כמו כן, לא נמצאה קורלציה (Mantel test) מובהקת בין המרחק הפיזי בין הקבוצות לבין המרחק הגנטי שביניהם (rxy=0.075; P=0.32; R2=0.0056; y=0.0001x + 0.0625). על פי מבחן לבדיקת השונות המולקולרית (AMOVA), רק 5% מהשונות הגנטית נבעה מהבדלים בין הקבוצות, שכל אחת מתחת לעץ אורן שרוף נפרד. 95% מהשונות הגנטית נבעה מהבדלים בין הפרטים ולא בין הקבוצות (טבלה 2).
טבלה 2. תוצאות מבחן לבדיקת השונות המולקולרית (AMOVA) הכוללת, וחלוקתה בין הקבוצות של הזריעים (כל אחת מתחת לעץ אורן שרוף נפרד), בין הפרטים בין הפרטים ובתוך הפרטים. P<0.01 מציין תוצאה מובהקת.
| מקור השונות | דרגות חופש | ס"ה הסטיות המרובעות | סטייה ממוצעת | שונות | % | F | P |
| בין הקבוצות | 19 | 127.345 | 6.702 | 0.153 | 5 | FST = 0.054 | 0.001 |
| בין הפרטים | 161 | 632.550 | 3.929 | 1.234 | 43 | FIS = 0.458 | 0.001 |
| בתוך הפרטים | 181 | 264.500 | 1.461 | 1.461 | 51 | FIT = 0.487 | 0.001 |
| ס"ה | 361 | 1024.395 | 2.848 | 100 |
סיווג לאשכולות (Cluster analysis UPGMA) על פי המרחק הגנטי (ההופכי של דמיון) של 78 זריעי אורן, שגדלו בקבוצות נפרדות מתחת ל 20 עצים שרופים גדולים שנה וחצי לאחר שריפה באתר לובים בכרמל לא הראה שהזריעים התקבצו לאשכולות בהתאם לקבוצות בהן הן גדלו (איור 6).
 איור 6. סיווג לאשכולות (Cluster analysis UPGMA) של 78 זריעי אורן, שגדלו בקבוצות מתחת לעצים שרופים גדולים שנה וחצי לאחר שריפה באתר לובים בכרמל על פי המרחק הגנטי (ההופכי של דמיון) ביניהם. כל קו מציין זריע אחד והמספר מימין את מספר הקבוצה בו הוא גדל. הקו האדום מדגים את המרחקים הגנטיים בין כל הזריעים בקבוצה 10 המפוזרים בין כל שאר הצברים ואינם יוצרים קבוצה אחת.
איור 6. סיווג לאשכולות (Cluster analysis UPGMA) של 78 זריעי אורן, שגדלו בקבוצות מתחת לעצים שרופים גדולים שנה וחצי לאחר שריפה באתר לובים בכרמל על פי המרחק הגנטי (ההופכי של דמיון) ביניהם. כל קו מציין זריע אחד והמספר מימין את מספר הקבוצה בו הוא גדל. הקו האדום מדגים את המרחקים הגנטיים בין כל הזריעים בקבוצה 10 המפוזרים בין כל שאר הצברים ואינם יוצרים קבוצה אחת.

דיון
דמוגרפיה והשפעת המרחק מהעץ השרוף
שלב מוקדם (שנתיים עד חמש שנים לאחר השריפה)
לאחר השריפה של 1989 בכרמל, נוצרו עיגולים מכוסים שכבה של אפר מתחת לצמרות השרופות של עצי אורן גדולים שנשרפו ומתו. שנתיים לאחר השריפה כמעט ולא נראו נבטי צמחים בעיגולי האפר, זאת לעומת הצפיפות הגדולה מאוד של נבטי אורנים, לוטם ומינים עשבוניים חד-שנתיים רבים שהיו רחוק מהעצים השרופים (Izhaki et al. 1992, Ne’eman et al. 1992). תוצאות המחקר מראות ששנתיים לאחר השריפה צפיפות נבטי האורנים הייתה נמוכה מאוד סמוך לגזעים השרופים וגבוהה מאוד רחוק מהם, אבל לא היה הבדל בגודלם. זאת למרות שמקורם של מרבית הנבטים הוא מזרעים שפוזרו מאצטרובלים אפילים מיד לאחר השריפה, שצפיפותם סמוך לעצים היא הגבוהה ביותר והיא יורדת עם המרחק מהעץ (Saracino et al. 2004). הסיבה לכך היא ה pH הגבוה בסיסי מאוד של האפר המעכב את הנביטה של צמחים רבים כולל זרעי אורן (Henig-Sever et al. 1995, Ne’eman 1997). האפר מורכב ממינרלים שמקורם בחומר הצמחי שנשרף, ולכן בשלב מאוחר יותר הוא מהווה דשן המעודד צמיחה של כל מי שהצליח לנבוט (Lamont et al. 1993). לכן, במשך השלוש השנים הבאות חלים שנויים יסודיים בגודל ובדגם המרחבי של זריעי האורנים. באתר שנבדק חמש שנים לאחר שריפה, צפיפות הנבטים הקרובים לגזעים השרופים, ששרידותם היית כ 50%, הייתה גבוהה מזו של המרוחקים שהתמותה בקרבם הייתה כ 75%. לתמותה הנמוכה יותר קרוב לגזעים מאשר במרחק מהם מספר גורמים אפשריים. א. תחרות תוך- ובין-מינית נמוכה שם יחסית לזו שבמרחק מהגזע (Ne’eman et al. 1992). ב. הנבטים הקרובים לגזעים נהנים מההזנה המינרלית המשופרת תודות לאפר (Kutiel and Naveh 1987). ג. תחרות נמוכה יותר עם המינים המתחדשים ווגטטיבית, שצפיפותם הייתה נמוכה יותר קרוב לגזעים השרופים. כתוצאה מסיבות אלה, זריעי האורן, שצמחו קרוב לגזעים השרופים, צמחו מהר יותר וגודלם חמש שנים לאחר השריפה היה כמעט פי 100 מאשר שנתיים לאחר השריפה, ואלו ששרדו רחוק צמחו רק פי 60.
שלב מאוחר (חמש עד עשרים שנים לאחר השריפה)
בשלב זה הייתה ירידה של 73% בצפיפות הנבטים הקרובים לגזע ושל 69% רחוק ממנו עד כי אחרי 20 שנים הצפיפות קרוב ורחוק מהגזע השרוף הייתה כעץ אחד למ"ר ולא היה יותר הבדל ביניהם. התמותה הגדולה קרוב נגרמה כנראה בשל ההצלה של עצים גדולים על קטנים מהם. בתקופה זו הביומסה של הזריעים הקרובים עלתה פי 168 וזו של הרחוקים פי 31 בלבד. בסוף התקופה, הזריעים הקרובים היו בעלי ביומסה גדולה פי 8.5 מאשר זו של הרחוקים מהגזע השרוף. זו כנראה תוצאה של הבדלים בתנאי מיקרו בית הגידול, ובמיוחד בעומק הקרקע באתר שגדל בו האורן הגדול השרוף. התוצאה היא שלמרות מות העץ הבוגר, אתר הגידול שלו הוא האתר המועדף להתחדשות הדור הבא לאחר שריפה. תופעה דומה שנמצאה גם במין אחר של אורן P. attenuata בקליפורניה (Keeley et al. 1999).
מודל מרחבי
המודל הקלסי של ג'אנסן (1970 Janzen) מתאר גיוס והישרדות של צאצאים יחסית למרחקם מצמח האם. על פי המודל, צפיפות הנבטים יורדת עם המרחק אבל שרידותם עולה עם המרחק עד שמגיע לשיא במרחק מסוים, שממנו מתחילה השפעה של עץ שכן. המודל מתייחס למקרים בהם צמח האם חי, והמכניזם המוצע הוא הקטנת התחרות עם צמח האם ובין הצאצאים עצמם, וכן הקטנת לחץ של הרביוורים ומזיקים אחרים הנמשכים לצמח האם ופוגעים גם בצאצאים. מודל זה אינו מתאים לתיאור התחדשות אורנים לאחר שריפה מכיוון שהאורנים מתים בשריפה. על סמך התוצאות כאן ושל שילר וחב. (1997 Schiller et al), אנו מציעים מודל אחר לתיאור ההתחדשות של עצים שמתים בשריפה ומתחדשים אחריה מנביטת זרעים בלבד. המודל מתאר את המגמות של השינויים בצפיפות ובגודל (ביומסה) של זריעים הגדלים קרוב למיקום הצמח הבוגר שמת בשריפה, לעומת אלה הגדלים רחוק ממנו (איור א מודל).
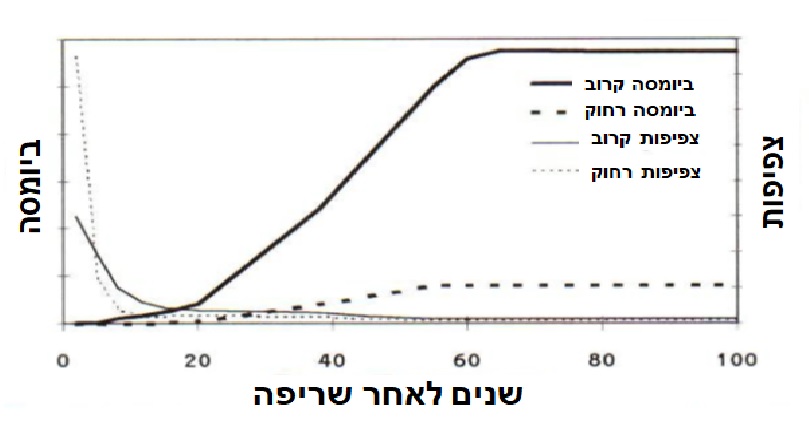
איור 7. מודל המתאר את השנויים החלים עם השנים לאחר שריפה בצפיפות של זריעי אורן הגדלים קרוב או רחוק מהזגע של אורן גדול שמת בשריפה (צפיפות קרוב וצפיפות רחוק) ובביומסה של זריעי אורן הגדלים קרוב או רחוק מהזגע של אורן גדול שמת בשריפה (ביומסה קרוב וביומסה רחוק).
בניגוד למודל של Janzen (1970) התנאים האופטימליים להישרדות והצלחה של הצאצאים לאחר שריפה הם ממש מתחת לצמרת השרופה. בתחילה צפיפות הנבטים גבוהה יותר במרחק מהעץ השרוף, אבל בשל תמותה גבוהה יותר, צפיפות הזריעים יורדת עד שהיא מגיע לרמה הדומה לזו הקיימת קרוב לעץ השרוף. אולם הנבטים המועטים קרוב לגזע צומחים במהירות והופכים לזריעים מבוססים שהם גדולים בהרבה מהמעטים ששורדים רחוק מהגזע. על פי תוצאות נמחקר, התהליך נמשך כ 40-20 שנים, שכן לא נמצא הבדל באחוז כיסוי הצמרות של עצי אורן בין יער שהתחדש במשך 40 שנים לאחר שריפה, לבין יער, סמוך בן כ 60, שלא נשרף. בניגוד לכך, גובה העצים ביער בן 60 שלא נשרף היה כפול מזה שביער שהתחדש לאחר שריפה (Schiller et al. 1997). קצב התחדשות דומה נצפה גם ביערות אורן ירושלים במערב אגן הים התיכון וצפון אפריקה (Trabaud et al.1985, Moravec 1990).
אנו מציעים שיש מספר תכונות אדפטיביות באורן ירושלים ובמינים דומים אחרים, המבטיחים שאתר שהיה מתאים לעץ גדול לצמוח ולהתרבות במשך עשרות שנים, ימשיך להיות מאוכלס על-ידי עץ אורן גם בדור המתחדש מזרעים לאחר שריפה, למרות שהעץ הבוגר מת בה (Ne’eman et al. 2004). חלק מתכונות אלה מאפיינות סינדרום של צמחים הגדלים במקומות בהם יש לעיתים מזומנות שריפות צמרות חזקות הגורמות לחילוף דורות ביער:ִ
- דליקות גבוהה של עלים וענפים בשל התכולה הגבוהה של שרפים ועלים מחטניים או קטנים בעלי שטח פנים גדול יחסית לנפחם. בנוסף, ענפים מתים ואצטרובלים ריקים נשארים על העץ ואינם נושרים כפי שקורה במרבית מיני העצים. תכונות אלה גורמות לדליקות הגבוהה של עצים ובמיוחד הזקנים שצברו הרבה ביומסה מתה (Keeley and Zedler 1998).
- הקליפה הדקה גורמת לתמותה של העץ בשל חימום ומוות של רקמת הקמביום הנמצאת בחלקו החיצוני של הגזע ואחראית על יצירת רקמות הובלה חדשות מידי שנה, וכניסת אויר לתאים המובילים מים (טרכאידות) המפסיקה את פעילותם. יצירת שריפה חזקה גורמת לירידה ביכולת ההתחדשות של צמחים צעירים המתחדשים ווגטטיבית. גורמת לתמותה של זרעים בשכבת הקרקע העליונה, עובדה המקנה יתרון יחסי למתחדשים מזרעים (Bond and Midgley 1995). אורן ירושלים הוא בעל אצטרובלים אפילים הנשמרים בצמרת העץ ומפזרים את זרעיהם בעיקר לאחר שריפה. מאגר זה מבטיח מלאי זרעים להתחדשות אחרי שריפת צמרות (Goubitz et al. 2004).
- הצבע הכהה של הזרעים, והעובדה שצידם האחד חום והשני שחור מסווים את הזרעים הנמצאים על הקרקע לאחר השריפה ובכך מפחיתים את שיעור הטריפה שלהם (Saracino et al. 2004, Lev-Yadun & Ne’eman 2013).
- שכבת האפר העבה מתחת לעצים גדולים שנשרפו היא בעל pH בסיסי מאוד המעכב נביטה ובכך מקטין את צפיפות הנבטים (Ne’eman et al. 1992), דבר הנראה תחילה כפוגע בהצלחת האורנים. אולם האפר מהווה הזנה מינרלית משופרת לפרטים שהצליחו לנבוט, גורם להישרדות גבוהה וצמיחה מהירה שלהם (Eshel et al. 2000).
סינדרום זה של תכונות מגדיל את ההסתברות שעץ אורן חדש יחליף באותו אתר את זה שמת בשריפה.
הרכב ומבנה גנטי
השפעה של שריפות על המבנה הגנטי של אוכלוסיות המתחדשות אחריה משתנה במערכות אקולוגיות שונות, וקשור לאקולוגיה, למנגנון הזיווג ולדמוגרפיה של אוכלוסיות אלה (Steinitz et al. 2012). למיטב ידיעתנו זו הפעם הראשונה שנערכת אנליזה גנטית לאוכלוסייה המתחדשת מזרעים לאחר שריפה. בניגוד לתחזית, לא נמצא מבנה מרחבי מסודר בקרב אוכלוסיית הנבטים. לא נמצאה קורלציה בין המרחק הפיזי למרחק הגנטי בין הפרטים, וניתוח השונית הגנטית הראה שמרביתה אינה בין קבוצת הנבטים שנבטו מתחת לעצים שונים, אל בין הפרטים בתוך כל קבוצה.
מרחק הפצת הזרעים האפקטיבי של אורן ירושלים והתבססות הצאצאים ללא שריפה נע בין 70-20 מ' מהעץ המפיץ, ומרבית הנבטים לא נמצאו סמוך לעץ האם (Troupin et al., 2006). ניתוח הורות של אורנים צעירים, הראה הפצת זרעים אפקטיבית לטווח של 492-6 מ' עם מרחק ממוצע של 42 מ' מצמח האם (Steinitz et al., 2011). הפצה יעילה לטווח כזה, העולה על המרחק שבין עצים בוגרים ביער אורנים טבעי מונעת יצירת דגם גנטי כתמי באוכלוסייה המתחדשת מזרעים. עלים בצמרות עצים חיים, מקטינים את מהירות הרוח ומפריעים פיזית להפצת הזרעים. לכן טווח ההפצה לאחר שריפה צפוי להיות גדול יותר מאשר ביער חי ( Shohami & Nathan 2014) ומקטינה עוד יותר את הסיכויים ליצירת דגם גנטי מרחבי באוכלוסייה המתחדשת לאחר שריפה.
התוצאות מראות שלמרות שסביר מאוד שמרבית הזרעים מתחת לעץ השרוף הן של אותו עץ, לנבטים במרבית הקבוצות שנבדקו היו יותר מאם אחת.
מסקנות
לא רק עצים המתחדשים ווגטטיבית שומרים על מיקומם בבית הגידול לאחר שריפת צמרות. גם אורנים, ברושים ומיני עצים אחרים המתים בשריפה והם בעלי בנק זרעים האגור בצמרותיהם, מתחדשים טוב יותר דווקא באתרים בהם היו עצים בוגרים שמתו בשריפה. האתרים בהם צמחו עצים גדולים לפני שריפה הם המועדפים לנביטה, והתבססות של זריעים אחריה, וגדולים הסיכויים שאחד מהם יחליף את העץ שמת בשריפה. לכן המבנה המרחבי של חברת הצמחים המתחדשת לאחר שריפה דומה לזה של חברת הצמחים טרם השריפה. למרות זאת, מסתבר שמרבית קבוצות הנבטים הגדלות באתר של עץ גדול שנשרף ומת, היו צאצאים של יותר מאם אחת. זאת כנראה תוצאה כתוצאה מפיזור הזרעים ברוח, היעיל לטווח ארוך באורן ירושלים.
ספרות
פרבולוצקי א בונה ע זלוצקי מ ושות 2011 דו"ח וועדת ממשק היער ושיקום הכרמל. מוגש למשרד להגנת הסביבה.
קליאוט נ וקידר ג 1992 אירועי שריפות בישראל והגורם האנושי. אופקים בגיאוגרפיה 35/36: 23-24.
Auerbach M & Shmida A 1987 Spatial scale and the determinants of .plant species richness. Trends in Ecology and Evolution 2: 228-231
Barbéro M Loisel R Quézel P et al. 1998 Pines of the Mediterranean Basin In: Richardson DM (editor) Ecology and Biogeography of .Pinus. Cambridge University Press Cambridge pp 153-170
Biswell HH 1974 Effects of fire on chaparral. In: Koslowski TT & Ahlgren CE (editors) Fire and Ecosystems. Academic Press London pp 321-364
Bond WJ & Midgley JJ 1995 Kill thy neighbour: An individualistic argument for the evolution of flammability. Oikos 73: 79-85
Clarke PJ Lawes MJ Midgley et al. 2013 Resprouting as a key functional trait: how buds, protection and resources drive persistence after fire. New phytologist 197: 19-35
Davis WS Borchert MI & Odion DC 1989 Establishment of micro-scale vegetation patterns in maritime chaparral after fire. Vegetatio 84: 63-67
Enright NJ Fontaine JB Lamont BB et al. 2014 Resistance and resilience to changing climate and fire regime depend on plant functional traits. Journal of Ecology 102: 1572-1581
Eshel A Henig-Sever N & Ne’eman G 2000 Spatial variation of seedling distribution in an east Mediterranean pine woodland at the beginning of post-fire succession. Plant Ecology 148: 175-182
European Commission 2001 Forest Fires in Europe. Fire Campaign Report 2: 1-28
Frelich LE Calcote RR Davis MB & Pastor J 1993 Patch formation and maintenance in an old‐growth hemlock‐hardwood forest. Ecology, 74: 513-527
Goubitz S Werger MJA & Ne’eman G 2003 Germination response to fire-related factors of seeds from non-serotinous and serotinous cones. Plant Ecol. 169: 195–204
Goubitz S Nathan R Roitemberg R Shmida A & Ne’eman G 2004 Canopy seed bank structure in relation to: fire, tree size and density. Plant Ecology, 173: 191-201.
Henig-Sever N Eshel A & Ne’eman G 1995 pH and osmotic potential of pine ash as post-tire germination inhibitors. Physiologia Plantarum 96: 71-76
Horn HS 1975 Markoviian properties of forest succession. In: Cody ML & Diamond JM (editors) Ecology and Evolution of Communities. University Press Harvard pp 196-211
Izhaki I Lahav H and Ne’eman G 1992 Spatial distribution patterns of Rhus coriaria seedlings after lire in a Mediterranean pine forest. Acta Oecologica 13: 279 -289
Janzen DH 1970 Herbivores and the number of tree species in tropical forests. American Naturalist 104: 501-528
Keeley JE 1986 Resilience of Mediterranean shrub communities to fires. In: Dell B Hopkins AJM & Lamont BB (editors) Resilience in Mediterranean-type ecosystems. Dr. Junk Publications pp 95-113
Keeley JE & Fotheringham CJ 1997 Trace gas emissions and smoke-induced seed germination. Science, 276: 1248-1250
Keeley JE & Zedler PH 1998 Evolution of life histories in pines In: Richardson DM (editor) Ecology and Biogeography of Pinus. Cambridge Univ Press Cambridge pp 219-249
Keeley J E Ne’eman G & Fotheringham CJ 1999 Immaturity risk in a fire-dependent pine. Journal of Mediterranean Ecology 1: 41-48
Keeley JE Bond WJ Bradstock RA., & Pausas JG 2011 Fire in Mediterranean Ecosystems: Ecology, Evolution and Management. Cambridge University Press. Cambridge pp 515
Korol L Shklar G & Schiller G 2002 Diversity among circum-Mediterranean populations of Aleppo pine and differentiation from brutia pine in their isoenzymes: Additional results. Silvae Genetica 51: 35-41
Kutiel P & Naveh Z 1987 Soil properties beneath P. halepensis and Q. caliprinos trees on burned and unburned mixed forests on Mt. Carmel, Israel. Forest Ecology and Management 20: 11-24
Lamont BB Le Maitre DC Cowling RM & Enright NJ 1991 Canopy seed storage in woody plants. The Botanical Review, 57: 277-317
Lamont BB Witkowski ETF & Enright NJ 1993 Post-tire litter microsites: Safe for seeds, unsafe for seedlings. Ecology 74: 501-512
Lev-Yadun S & Ne’eman G 2013. Bimodal colour pattern of individual Pinus halepensis Mill. seeds: a new type of crypsis. Biological Journal of the Linnean Society 109: 271-27.
Malanson GP 1987 Diversity, stability and resilience: Effect of fire regimes. In: Trabaud L (editor) The Rote of Fire in Ecological Systems. SPB Academic Publications The Hague pp. 49-63
Mataix-Solera J Cerdà A Arcenegui V Jordán A & Zavala LM 2011 Fire effects on soil aggregation: a review. Earth-Science Reviews 109: 44-60
Moravec J 1990 Regeneration of NW Africa Pinus halepensis forests following fire. Vegetatio 87: 29 36
Moreno JM & Oechel WC 1991 Fire intensity effects on germination of shrubs and herbs in southern California chaparral. Ecology 72: 1993 2004
Nathan R Safriel UN Noy-Meir I & Schiller G 1999 Seed release without fire in Pinus halepensis, a Mediterranean serotinous wind-dispersed tree. Journal of Ecology 87: 659-669
Naveh Z 1994 The Role of Fire in Mediterranean-type Ecosystems. Ecological Studies 107: 163-185.
Naveh Z 1989 Fire in the Mediterranean landscape, an ecological perspective. In: Goldammer JG & Jenkins MJ (editors) Fire in Ecosystem Dynamics. SPB Academic Publishers The Hague pp. I-20.
Ne’eman G 1994 The effect of thinning out on the survival and development of Pinus and Cisius seedlings after fire on Mt. Carmel, Israel. In: Vieaas DX (editor) Proceedings of Second International Conference on Fire Research Coimbra Portugal pp 1021-1031
Ne’eman G 1997 Regeneration of natural pine forest – review of work done after the 1989 fire in Mount Carmel. Israel. International Journal of Wildland Fire 7: 295 306
Ne’eman G & Izhaki I 1999 The effect of stand age and microhabitat on soil seed banks in Mediterranean Aleppo pine forests after fire. Plant Ecology 144: 115-125
Ne’eman G & Trabaud L (editors) 2000 Ecology, Biogeography and Management of Pinus halepensis and P. brutia Mediterranean Pine Forest Ecosystems.: Buckhuys Publishers Leiden
Ne’eman G Lahav H & Izhaki I 1992 Spatial pattern of seedlings one year after fire in a Mediterranean pine forest. Oecologia 91: 365-370
Ne’eman G Goubitz S & Nathan R 2004. Reproductive traits of Pinus halepensis in the light of fire – a critical review. Plant Ecology 171: 69–79
Pausas JG 2015 Evolutionary fire ecology: lessons learned from pines. Trends in Plant Science 20: 318–324
Pausas JG & Keeley JE 2017 Epicormic resprouting in fire-prone ecosystems. Trends in Plant Science 22: 1008-1015
Pausas JG Bradstock RA Keith DA & Keeley JE 2004 Plant functional traits in relation to fire in crown‐fire ecosystems. Ecology 85: 1085-1100
Quézel P 2000 Taxanomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In: Ne’eman G & Trabaud L (editors) Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin. Backhuys Publishers Leiden pp 1-12
Rice SK 1993 Vegetation establishment in post-fire Adenostonia chaparral in relation to fine scale pattern in fire intensity and soil nutrients. Journal Vegetation Science 4: 115-124
Saracino A & Leone V 1993 Natural regeneration 2 and 4 years after fire of Pinus halepensis Mill. in dunal environment. In: Trabaud L & Prodon R (editors) Fire in Mediterranean ecosystems. Commission of the European Community. Brussels-Luxembourg pp141-150
Saracino A D’Alessandro CM & Borghetti M 2004 Seed colour and post-fire bird predation in a Mediterranean pine forest. Acta Oecologica, 26: 191-196.
Schiller G Ne’eman G & Korol L 1997 Post-fire vegetation dynamics in a native Pinus halepensis Mill. forest on Mt. Carmel Israel. Israel Journal of Plant Sciences 45: 297-308
Shmida A & Whittaker RH 1981 Patterns and biological microsite in two shrub communities, Southern California. Ecology 62: 234 251
Shohami D & Nathan R 2014 Fire-induced population reduction and landscape opening increases geneflow via pollen dispersal in Pinus halepensis. Molecular Ecology 23: 70-81
Steinitz O Troupin D Vendramin GG & Nathan R 2011 Genetic evidence for a Janzen-Connell recruitment pattern in reproductive offspring of Pinus halepensis trees. Molecular Ecology 20: 4152-4164
Steinitz O Shohami D Ben-Shlomo R & Nathan R 2012 Genetic consequences of fire to natural populations. Israel Journal of Ecoloy and Evolution 58: 205-220
Thanos CA Georghiou K Kadis C & Pantazi C 1992 Cistaceae: A plant family with hardseeds. Israel Journal of Botany 41: 251-263
Trabaud L 1994 Post tire community dynamics in the Mediterranean basin. In: Moreno JM & Oechel WC (editors) The Role of Fire in Mediterranean Ecosystems. Springer New York pp 1-15
Trabaud L & Oustric J 1989 Influence du leu sur la germination de semences de quatre espcces ligneuses mediterranees a reproduction sexuee obligatoire. Seed Science and Technology 17: 589-599
Trabaud L Michels C & Grossman J 1985 The recovery of burned Pinus halepensis Mill. forests II Pine reconstituton after wild-fire. Forest Ecology and Management 13: 167-179
Turner MG & Romme WH 1994 Landscape dynamics in crown fire ecosystems. Landscape Ecology 9: 59-79
המאמר מבוסס על המאמרים הבאים:
Ne’eman G & Izhaki I 1998 Stability of pre- and post-fire spatia structure of pine trees in Aleppo pine forest. Ecography 21: 535-542
Gershberg A Ne’eman G & Ben-Shlomo R 2016 Genetic structure of a naturally regenerating post-fire seedling population: Pinus halepensis as a case study. Frontiers in Plant Science 7: 549
================
כל הזכויות שמורות ל"כלנית" ©
לציטוט: נאמן ג 2021 הפיזור המרחבי וההרכב הגנטי של עצי אורן ירושלים טבעיים לפני ואחרי שריפה, כתב-עת "כלנית", מספר 8.
================










לפוסט הזה יש 2 תגובות
ציינת רק את מספר האמהות שזוהו בכל קבוצת זריעים. האם ניתן לקבוע את אחוז הצאצאים של כל אחת מהאמהות בקבוצה?
האם יש ניתן לאשר או לשלול את ההשערה שרוב הזריעים באותו מקום מקורם באם אחת ואילו הנוכחות של צאצאים מאמהות אחרות מועטת?
בשל כשל במיצוי DNA מהעצים המתים לא יכולנו לקבוע מי וכמה מהנבטים הם אכן צאצאי העץ המת.