

גידי נאמן החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים, gneeman@gmail.com
רנה נאמן מושב אמירים rina.nem@gmail.com
צמחים משתמשים באותות חזותיים (ראייה) וכימיים (ריח) על מנת למשוך ולכוון את המאביקים לפרחיהם. הראייה, כושר ההפרדה, יכולת הזיהוי, מרחק הגילוי המרבי וזווית הראיה המזערית של מטרות על-ידי דבורים נבדקו לראשונה במחקר זה בפרחים אמיתיים ולא במטרות עגולות מנייר צבעוני. במחקר זה השתמשנו במבוך רדיאלי בעל שש זרועות לקביעה של מרחק הגילוי המרבי וזווית הראייה המזערית של 12 סוגי פרחים טבעיים על-ידי פועלות של בומבוס האדמה (Bombus terrestris), וניתחנו את הגורמים המשפיעים עליו. נמצא שמרחק הגילוי אינו קשור לקוטר הפרח, אבל יש לו קשר חיובי לשטח הפרח, וקשר שלילי לאורך קו המתאר שלו. ככל שהיחס בין שטח הפרח לאורך קו המתאר קטן יותר ולפרח צורה גזורה יותר, מרחק הגילוי שלו קטן יותר. מרחק הגילוי של פרחים מלאים ועגולים גדול יותר מאשר זה של פרחים בקוטר דומה אך גזורים. נמצאה נוסחה אמפירית, המאפשרת לחזות את מרחק הגילוי המירבי של כל פרח על-פי משתנים אלה. קביעת מרחק הגילוי והגורמים המשפיעים עליו, חשובה לחקר התנהגות איסוף המזון של דבורים, האבולוציה של תכונות הפרחים, ולאימות של מודלים נוירופיסיולוגיים המסבירים את מנגנון הראייה בדבורים.
מאמר זה מבוסס על המאמר:
Ne’eman G and Ne’eman R 2016 Factors determining visual detection distance to real flowers by bumble bees. Journal of Pollination Ecology 20: 1-12. http://www.pollinationecology.org/index.php?journal=jpe&page=article&op=view&path%5B%5D=416&path%5B%5D=143.
מבוא
מאביקים מבקרים בפרחים על מנת לאסוף גמול, בעיקר צוף ואבקה (Heinrich, 1975). הצמחים משתמשים בריח ובסימנים חזותיים כמו גודל, צורה וצבע כפרסומת על מנת למשוך ולכוון אותם לפרחיהם (Chittka & Raine, 2006; Dafni et al., 1997; Menzel & Shmida, 1993; Schaefer et al., 2004). כבר במאה ה 18 זוהתה החשיבות של גודל הפרח וצורתו "במשיכה מרחוק של עין החרקים" (Sprengel, 1793, in Hasse, 1996). הזיהוי של צורה וצבע על-ידי דבורים מבוסס על יכולת הראיה והלימוד של הדבורים, כפי שנמצא בעבודותיו של חתן פרס הנובל קרל פון פריש (von Frisch, 1914) ואחרים (Hertz, 1929) . Kugler (1934) היה הראשון שקבע את המרחק ממנו גילו דבורי בומבוס דגמי פרחים על פי המקום בו שינו את כיוון מעופן ישירות לדגמי הפרחים, והראה שדגמים גדולים מתגלים ממרחק גדול יותר מאשר פרחים קטנים.
בהתאמה לעקרון 'אמת בפרסום' (Armbruster et al,. 2005; Keasar et al., 2006), פרחים גדולים המתגלים ממרחק גדול יותר (Chittka & Spaethe, 2007), מכילים בדרך כלל יותר צוף או אבקה מאשר פרחים קטנים (Blarer et al,. 2002; Fenster et al., 2006; Herrera 2005; Martin 2004). מאביקים מעדיפים בדרך כלל פרחים גדולים על פני קטנים, מבקרים בהם בתדירות גבוהה יותר ומאביקים אותם ביעילות רבה יותר (Conner & Rush, 1966; Elle & Carney, 2003; Fenster et al,. 2006). לכן, הכשירות של צמחים בעלי פרחים גדולים, המתגלים ממרחק גדול יותר על-ידי מאביקיהם, אמורה להיות גדולה יותר מאשר של צמחים בעלי פרחים קטנים, וכתוצאה מכך מאביקים יכולים להפעיל לחץ ברירתי להגדלת הפרחים (Galen & Newport, 1987).
אולם בנוסף ללחץ ברירתי המופעל על-ידי המאביקים, גודל הפרח מושפע גם ממנגנון הזיווג של הצמח (Goodwillie et al., 2010), מהרביוורים אוכלי פרחים ועלים (Galen, 1999), ושלל גורמים אביוטיים וביוטיים נוספים (Krizek & Anderson, 2013). הבדלים בגודל ובתכונות שונות של פרחים, בין אוכלוסיות מבודדות של מין מסוים, עשויות להוביל להתמיינות וליצירה של מינים חדשים (Krizek & Anderson, 2013).
(Giurfa et al. (1996 קבעו את זווית הראיה המזערית הדרושה לפועלות של דבורי דבש (Apis mellifera) על מנת לראות דיסקיות נייר צבעוניות, על-ידי שינויים בקוטר של דיסקיות הנייר העגולות. הם מצאו שזווית הראיה המזערית היא קבועה ללא תלות בקוטר המטרה או במרחק שלה, אך תלויה בצבעה. זווית זו היא 5º עבור מטרות בעלות ניגודיות באורך גל ירוק, בינן לבין הרקע, ו 15º עבור מטרות דומות אבל ללא ניגודיות כזו (Giurfa et al. 1996). כלומר, שמרחק הגילוי של מטרות בעלות ניגודיות בירוק בינן לבין הרקע – גדול יותר מאשר זה של מטרות דומות חסרות ניגודיות בירוק, זאת, ללא תלות בניגודיות הצבע בכל אורכי הגל האחרים אותם רואות הדבורים (Giurfa et al. 1996; Giurfa & Vorobyev 1997; Lehrer & Bischof 1995). בהמשך לממצאים אלה, נערכו הדמייות מחשב לקביעת המרחקים ממנו דבורי דבש יכולות לגלות פרחים על סמך הקוטר והצבע שלהם בלבד (Giurfa & Vorobyev, 1997; Menzel et al., 1997; Vorobyev et al., 1997).
בשונה מדבורי דבש, דבורי בומבוס מסוגלות לגלות מטרות עגולות מלאות בעלות ניגודיות בירוק החל מזווית ראיה של 2.3°, ושל מטרות חסרות ניגודיות כזו החל מזווית של 2.7° (Dyer et al. 2008). לאחרונה נקבעו זוויות הראיה המזעריות גם עבור דבורים חסרות עוקץ מאוסטרליה, שהן 9.3° עבור מטרות בעלות ניגודיות בירוק, ו 9.5° עבור מטרות חסרות ניגודיות בירוק (Dyer et al. 2016). הבדלים אלה נובעים, כנראה, מכך שרק לדבורי דבש יש את היכולת להעביר מידע על המרחק והכיוון של מקורות המזון (על-ידי 'ריקוד הדבורים'), ולכן לדבורי בומבוס ודבורים טרופיות חסרות עוקץ נדרשת יכולת טובה יותר לגילוי של פרחים קטנים חסרי ניגודיות בירוק (Dyer et al. 2008; Bukovac et al. 2013; Dyer et al. 2016).
מחקרים רבים על ראיית דבורים התרכזו במנגנונים נוירוביולוגיים, בתפיסה (פרספציה), ביכולת הלמידה ובזיכרון של דבורי דבש ודבורי בומבוס (Chittka & Raine, 2006; Menzel & Giurfa, 2006; Srinivasan, 2010; Avarguès-Weber et al., 2011; Avarguès-Weber & Giurf, 2014; Hempel de Ibarra et al., 2014; Hempel de Ibarra et al., 2015). התפקיד של הגודל הצורה והדגם בפרסומת של פרחים, וקליטתם על-ידי המאביקים, נחקרו רבות ונסקרו במאמרים רבים (Lehrer & Bischof 1995; Dafni and Kevan 1997; Dafni et al. 1997; Neal et al. 1998; Zhang et al. 2004; Kaczorowski et al. 2012; Avarguès-Weber et al. 2011; Hempel de Ibarra et al. 2015). בדיקה מדוקדקת הראתה שמרבית החוקרים השתמשו במטרות נייר עגולות, ולא בפרחים אמיתיים. הם השתמשו בקוטר ובצבע של מטרות הנייר לקביעת זווית הראיה המזערית ומרחק הגילוי שלהן, והקישו מכך על תהליך הגילוי של פרחים על-ידי הדבורים. אולם מרבית הפרחים אינם עיגול מלא אלא הם בעלי צורה רדיאלית גזורה או צורה לא רגולרית בעלת סימטריה דו-צדדית.
אם הקוטר הוא הגורם העיקרי הקובע את מרחק הגילוי של פרח על-ידי דבורים, הרי צמחים יכולים לייצר פרחים גזורים בעלי ביומסה קטנה ועלות נמוכה, במקום פרחים מלאים בעלי ביומסה ועלות ייצור גדולות יותר, זאת מבלי להקטין את יעילות הפרסומת של הפרחים. את החיסכון הם היו יכולים להשקיע בצמיחה או בייצור זרעים ולהגדיל בכך את כשירותם. לכן, במחקר קודם, בדקנו את מרחק הגילוי של מטרות נייר צבעוני בעל ניגודיות בירוק, שהיו בצורות שונות, דמויות פרחים גזורים ובעלות קוטר שונה (Ne’eman and Kevan, 2001). מצאנו שהקוטר לא קבע את מרחק הגילוי, אבל ככל ששטח המטרה היה גדול יותר, גם מרחק הגילוי היה ארוך יותר. לעומת זאת, ככל שקו המתאר היה ארוך יותר, מרחק הגילוי היה קצר יותר (Ne’eman and Kevan, 2001). כלומר, בניגוד למצופה, על מנת לגלות דגמים של פרחים גזורים, דבורת דבש חייבת להתקרב יותר מאשר אל דגם של עיגול מלא. לכן לא ניתן להשתמש בקוטר הפרח על מנת להעריך את מרחק הגילוי של פרחים על-ידי דבורים.
אורך קו המתאר ידוע כגורם חשוב המשפיע על יכולת ההבחנה וההתנהגות של דבורים. כבר (von Frisch (1914 מצא שדבורים מסוגלות ללמוד להבחין בין צורות הנבדלות באורך קו המתאר ומידת הגזירות שלהן, אבל לא בין צורות מלאות של משולש, ריבוע או עיגול. במחקר של מידת הדמיון בהן דבורים תופסות מטרות בעלות צורות שונות, נמצא ששטח המטרה וצפיפות קווי המתאר שלה (היחס בין אורך קו המתאר לשטח העיגול המכיל את המטרה), הסביר בצורה הטבה ביותר את מידת הדמיון של צורות בעיני הדבורים (Anderson, 1977).
דבורי דבש מסוגלות להבחין ולהפריד בין דגמים וללמוד הבדלים מורכבים הרבה יותר מאשר נמצא בתחילה (Avarguès-Weber et al., 2011; Srinivasan, 2010). דבורים מסוגלות ללמוד ולקבץ מטרות על פי קטגוריות שונות כמו: פרחים דמויי כוכב בצבעים שונים, תמונות של פרחים עגולים בצבעים שונים, תמונות של גבעולי צמחים בצורות שונות וגם תמונות נוף (Zhang et al,. 2004). נמצא שדבורי דבש אפילו מסוגלות ללמוד את המאפיינים של סגנון הציור של ציירים ולהבדיל בין ציורים של Monet לבין ציורים של Picasso (Wu et al. 2013). נמצא גם שדבורי דבש מסוגלות ללמוד יחסי גודל ולבחור במטרות גדולות או קטנות בהתאם למיקום הגמול וליישם את החוק שנלמד לגבי מטרות גדולות וקטנות אך בצבעים וצורות שונים (Avarguès-Weber et al., 2014).
למרות המחקרים הרבים מאוד על יכולת הראיה, ההבחנה והלמידה של דבורים תוך שימוש במטרות מלאכותיות וסימולציות מחשב, טרם נקבע מרחק הגילוי המירבי של פרחים טבעיים על-ידי דבורים.
מטרת מחקר זה היא לקבוע לראשונה את מרחק הגילוי המירבי של פרחים אמיתיים על-ידי פועלות של דבורי בומבוס. בנוסף אנו מבקשים לחקור את הגורמים המשפיעים על מרחק הגילוי ולבנות מודל שבעזרתו ניתן יהיה לחזות את מרחק הגילוי על סמך צורת הפרח וצבעו. זאת מכיוון שלמרחק הגילוי של פרחים יש החשיבות רבה בקביעת התנהגות איסוף המזון של דבורים ולכן גם השפעה על האבולוציה של תכונות הפרחים, זאת באמצעות הברירה הטבעית שמפעילים המאביקים על תכונות הפרחים.
חומרים ושיטות
המבוך
בעקבות חוקרים אחרים (Lehrer et al., 1995; Simonds & Plowright, 2004; Plowright et al., 2011; Thompson & Plowright, 2014) השתמשנו מבוך רדיאלי בקוטר 100 ס"מ בעל שש זרועות (איור 1א). השתמשנו במבוך זה משום שמספר הניסיונות הדרוש על מנת לקבל תשובה מובהקת מבחינה סטטיסטית במבוך זה נמוך ממספר הניסיונות הדרוש במבוך דמוי Y (ראו Bukovac et al., 2016).
מושבה של דבורי בומבוס חוברה במעבר פרספקס (Plexiglas®) שקוף אל חור הכניסה שהיה במרכז רצפת המבוך. במעבר הותקן שער שאיפשר זיהוי של מספר הדבורה והכנסת הדבורים אחת אחר השנייה למבוך. מסביב לפתח הכניסה, ובגבול הכניסה לזרועות המבוך, סומן עיגול צהוב בקוטר של 10 ס"מ, שסימן את גבולות ' תחום ההחלטה'. לכל דבורה שהגיחה למבוך, חצתה את הקו, ונכנסה לזרוע עם פרח גומל נרשמה הצלחה. לדבורה שנכנסה תחילה לזרוע ללא פרח גומל נרשם כישלון. הקו שימש גם למדידת המרחק אל פרח המטרה. כל זרוע של המבוך הייתה ברוחב של חמישה ס"מ ליד הקו הצהוב ו 10ס"מ בקצה המרוחק שלה. גובה המבוך היה 20 ס"מ והוא היה מכוסה במכסה שקוף (Plexiglas®) החדיר לקרינה אולטרה סגולה (UV). המבוך היה בצבע ירוק כהה ורק באחת משש זרועותיו הוצג במאונך פרח שגבעולו היה טבול במים מעבר למחיצה שאליה היה הפרח מחובר (איור 1ב). הפרח הוצג באופן שהדבורה ראתה אותו באופן חזיתי (en-face). המחיצה הייתה ניידת כך שניתן היה לשנות את המרחק של הפרח מקו ההחלטה הצהוב. בבסיס המחיצה היה כלי מכוסה עם תמיסת סוכר בריכוז 50% (סמ"ק/גר'). בניסוי מוקדם נמצא שהדבורים לא ראו את הכלי שהכיל את הגמול ממרחק של 15 ס"מ.
בשימוש בפרחים טבעיים במבוך חייבים להקפיד על הפרדת הגירוי החזותי (ראיה) מהכימי (ריח). מכיוון שהמבוך הוא חלל סגור אחד, ומיקום הפרחים שונה באקראי בין הזרועות, ניתן להניח שריח הפרחים היה אחיד בכל המבוך. למרות זאת הכנסנו לכל זרוע של המבוך פרחים טריים שהיו מוסתרים מעין הדבורים אך פתוחים לחלל המבוך (איור 1א).
המבוך הואר בנורות המספקות את כל אורכי הגל (TMS 24F lamps with 4.3 kHz ballasts; (Philips, Eindhoven, The Netherlands).

הפרחים
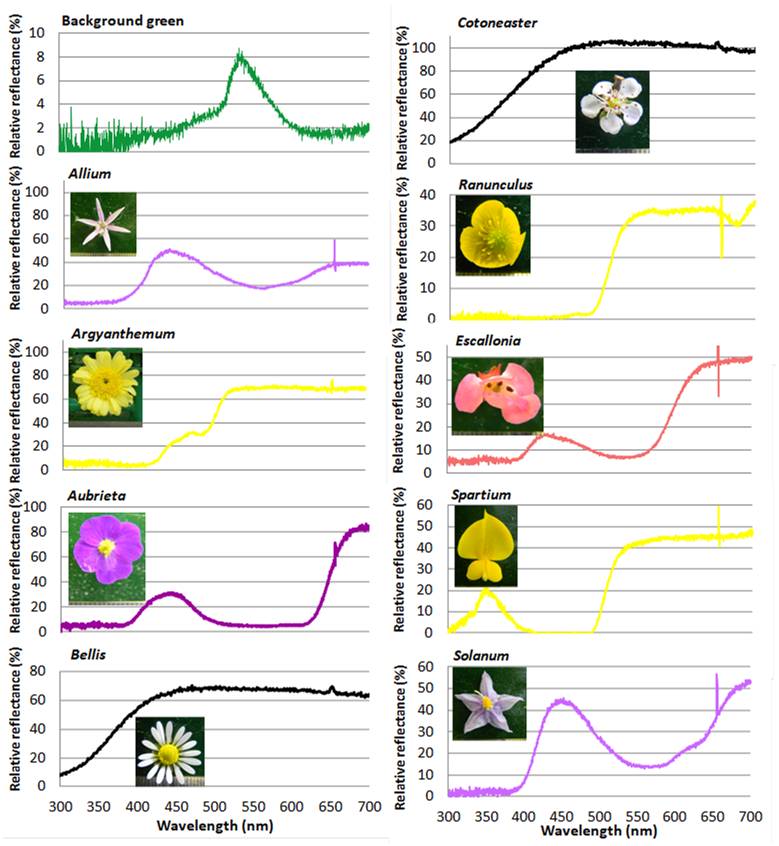
השתמשנו בפרחים של תשעה מיני צמחים (טבלה 1) שנקטפו מדי בוקר (אפריל-מאי 2012) בגנים של Queen Mary University of London וב Victoria Park הסמוך. הפרחים הוצגו במבוך במאונך, באופן שהדבורים ראו אותם באופן חזיתי (en-face). פרחי חיננית (Bellis perennis) ממשפחת המורכבים הוצגו כמות שהם, ובהמשך לאחר שנתלשו מהם פרחים לשוניים ליצירת דמות של פרחים בעלי 16 ו-8 עלי כותרת. פרחים של אחירותם החורש (Spartium junceum ) הוצגו בפרופיל ובאופן חזיתי. בסך הכול השתמשנו ב 12 פרחים שנבדלו בתכונות הבאות (טבלה 1): (1) קוטר המעגל המקיף -את הפרח, (2) שטח עלי הכותרת הצבעוניים, (3) קו המתאר (contour line, perimeter) של הפרח. (4) מידת הניגודיות ברצפטור של אורך הגל הירוק בין צבע הפרח לבין הצבע הירוק של המבוך. התכונות המורפולוגיות נמדדו בעזרת התכנה לניתוח תמונות ('CellProfiler 2.0' Massachusetts Technological institute), מתמונות מכוילות של הפרחים.
טבלה 1. השמות המדעיים של מיני הצמחים, שם הסוג בעברית, השטח, הקוטר, אורך קו המתאר הממוצע, והניגודיות בירוק בין הפרח לרקע של המבוך. המספר הצמוד לשם הצמח מציין את מספר עלי הכותרת (או הפרחים הלשוניים בקרקפת של מורכבים). השמות העבריים שלצמחי תרבות (בישראל) על פי טבלת השמות באתר משרד החקלאות 2011.
|
מין הצמח
|
שם עברי של הסוג | שטח (ממ"ר) | קוטר (מ"מ) | קו מתאר (מ"מ) | ניגודיות בירוק |
|
Allium cristophii |
שום | 111 | 17 | 128 | 0.379 |
|
Aubrieta deltoidea |
אובריטה | 314 | 20 | 74 | 0.221 |
|
Argyanthemum frutescens |
חרצית שיחית | 606 | 30 | 234 | 0.451 |
|
Bellis perennis |
חיננית | 112 | 12 | 47 | 0.499 |
|
Bellis perennis 8 |
חיננית | 134 | 16 | 142 | 0.499 |
|
Bellis perennis 16 |
חיננית | 242 | 22 | 259 | 0.499 |
|
Cotoneaster microphyllus |
חבושית | 105 | 12 | 61 | 0.463 |
|
Escallonia macrantha |
אסקולינה | 151 | 15 | 63 | 0.255 |
|
Ranunculus bulbosus |
נורית | 174 | 15 | 55 | 0.468 |
|
Solanum crispum sta |
סולנום | 190 | 21 | 126 | 0.427 |
|
Spartium junceum |
אחירותם (חזית) | 510 | 26 | 117 | 0.379 |
|
Spartium junceum |
אחירותם (פרופיל) | 288 | 25 | 113 | 0.379 |
ניתוח צבע הפרחים
עקומת ההחזר הספקטרלית של הפרחים ושל צבע הרקע של המבוך נבדקו בספקטרופוטומטר (Ocean Optics, Dunedin, FL, S2000 with a deuterium-halogen light source). משתני הצבע (העירור היחסי של הרצפטורים של הדבורה בירוק כחול ואולטרה סגול, וכן ניגודיות הצבע והניגודיות בירוק שבין הפרח לרקע חושבו על פי (Backhaus (1992, תוך שימוש במשושה הצבעים, המדמה את ראיית הצבע על-ידי הדבורה ומאפשר הערכת האפשרות של הדבורה להפריד בין צבע מסוים והרקע, ובין צבעים שונים (Chittka, 1992, Dyer & Chittka, 2004). בפרחים בעלי שני צבעים, בהם המרכז שונה בצבעו משאר הפרח, נמדד רק הצבע העיקרי של הפרח. כמו כן לא יכולנו למדוד את צבעם השונה של נתיבי הצוף בפרחים בהם הם נמצאים.
תמונות הפרחים, ועקומת החזר הצבע היחסית שלהם מוצגות באיור 2. המיקום של כל הפרחים הנבדקים במשושה הצבעים המותאם לדבורי בומבוס, מוצגים באיור 3. מרכז המשושה מייצג את הרקע (צבע המבוך) והמרחק של כל פרח ממרכז המשושה המייצג את ניגודיות הצבע יחסית לרקע (Chittka 1992).

הדבורים, ומהלך הניסויים
מושבה של דבורי בומבוס האדמה (Bombus terrestris) נקנתה מ Syngenta Bioline Bees בהולנד. הדבורים היו בקופסאות חשוכות עד לתחילת הניסוי. תחילה הורשו הדבורים להיכנס ולהתאקלם למבוך. בזמן זה, הן הורגלו להיזון ממתקן ההזנה שהוצב למרגלות המחיצה הניידת שהייתה סמוך מאוד לקו ההחלטה הצהוב, ועליה עיגול מטרה צהוב בקוטר חמישה ס"מ. דבורים שמצאו את מתקן ההזנה בקלות, נלכדו וסומנו במדבקות ממוספרות על הצד הגבי של החזה. הדבורים הממוספרות אומנו להיזון במתקן ההזנה שהיה למרגלות המחיצה הניידת, במרחק של חמישה ס"מ מקו ההחלטה ומעליו פרח אמיתי. בכל מחזור אימון ומבחן עבדנו עם שבע דבורים שרק אחת מהן הורשתה להיכנס בכל פעם למבוך. זאת על מנת שהדבורים לא תלמדנה את מיקום הגמול. לאחר סיום הביקורים של מחזור הדבורים, הוחלף מיקום הזרוע הגומלת על פי טבלה של מספרים אקראיים. לאחר שלושה מחזורי תרגול מוצלחים, נבדקו הדבורים לגבי פרחי מטרה שהוצבו במרחק של 15 ס"מ, לאחר מכן הוגדל המרחק בהדרגה לקביעת מרחק הגילוי המרבי. כל הדבורים שנבדקו היו אחיות מאותה מושבה.
כל דבורה שנכנסה למבוך נצפתה עד שמצאה את מתקן ההזנה, ניזונה ושבה לכוורת. לכל דבורה שנכנסה תחילה לזרוע הגומלת, שבה הוצג הפרח, נרשמה הצלחה ולדבורה שנכנסה תחילה לזרוע לא גומלת נרשם כישלון. כל הדבורים, גם אלה שנכשלו, הושארו במבוך עד שמצאו וניזונו במתקן לפני שובן לכוורת. כאמור, לאחר כל ביקור הוחלף מיקום הפרח והגמול באקראי. לאחר סיום מחזור הבדיקות במרחק מסוים הוגדל המרחק ב 10 ס"מ, עד שמספר ההצלחות לא היה שונה מאקראי, סימן לכך שממרחק זה הדבורה לא ראתה את הפרח. לאחר מכן צומצם המרחק בחמישה ס"מ והדבורה נבדקה מחדש. מרחק הגילוי המרבי נקבע כממוצע בין המרחק המרבי ממנו ראתה הדבורה את הפרח, לזה שממנו היא לא ראתה אותו. לכל סוג פרח נתון, נבדק מרחק הגילוי המרבי של חמש עד תשע דבורים שונות, שהיו אחיות מאותה כוורת. לאחר סיום הבדיקה של כל סוג פרח הורשו הדבורים של הכוורת להיכנס באופן חופשי למבוך, ודבורים נוספות מוספרו לקראת המבחן הבא.
זווית הראיה המזערית, שממנה מסוגלת הדבורה לגלות כל טיפוס פרח חושבה מהקוטר (D) הממוצע של הפרח ומרחק הגילוי המירבי (MDD) שלו:
זווית הראיה המזערית = (2arctan (D / 2MDD
ניתוח הנתונים
השתמשנו במבחן בינומיאלי (בחירה בין שתי אפשרויות: הצלחה או כישלון) לבדוק האם מספר ההצלחות של דבורה בגילוי המטרה מתוך כלל מספר הנסיונות, יכול להיות תוצאה של בחירה אקראית. ההסתברות של בחירה נכונה באקראי של אחת מתוך שש אפשרויות היא 0.17, על מנת לנקוט זהירות יתר בחרנו הסתברות של 0.2. כל דבורה נבדקה לפחות חמש פעמים עד שהתקבלה תוצאה מובהקת ועד 12 פעמים. על מנת להגיע למסקנה שדבורה ראתה את פרח המטרה ממרחק נתון (בהסתברות של 95%) היה על הדבורה להצליח ב 4/5, 4/6, 4/7, 5/9, 5/10, 5/11, או 6/12 בחירות. המרחק שממנו ערך ההסתברות היה גבוה מ P=0.05, כלומר מספר הבחירות הנכונות לא היה שונה באופן מובהק מבחירה אקראית, נקבע כמרחק הגילוי המרבי והוא שימש לחישוב זווית הראיה המזערית.
השתמשנו ברגרסיה לינארית על מנת לבנות מודל שיאפשר חיזוי של מרחק הגילוי המרבי של פרח נתון (משתנה תלוי) על פי תכונות הפרח הבאות (משתנים בלתי תלויים): (1) קוטר של המעגל המקיף את הפרח, (2) שטח עלי הכותרת הצבעוניים, (3) קו המתאר (contour line, perimeter) של הפרח. (4) מידת הניגודיות של הרצפטור באורך הגל הירוק בין צבע עלי הכותרת לבין הצבע הירוק של המבוך.את ההתפלגות הנורמלית של הנתונים בדקנו במבחן Kolmogorov-Smirnov. כל הניתוחים הסטטיסטיים בוצעו ב (SPSS 19 (IBM Corp., 2010.
תוצאות
הנתונים המורפולוגיים של כל הפרחים שנבדקו מוצגים בטבלה 1, עקומת החזרי הצבע היחסית מוצגת באיור 2, ומיקומם במשושה הצבעים של דבורי בומבוס באיור 3.
הקוטר הממוצע של 12 הפרחים שנבדקו (±סטיית תקן.) היה 1.9±0.6 ס"מ. טווח מרחק הגילוי המירבי היה בין 22 ל 38.8 ס"מ, כאשר הממוצע (±סטיית תקן.) היה 27.6±5.2 ס"מ, וההבדלים במרחק הגילוי של הפרחים השונים היו מובהקים (ANOVA, F11,63=6.458, P<0.001) (איור 4A).
השתמשנו ברגרסיה לינארית על מנת לגלות את המודל המתאים ביותר לניבוי מרחק הגילוי המירבי על סמך התכונות המורפולוגיות והצבעים של הפרחים. התוצאות מראות שמרחק הגילוי המירבי לא יכול להיות מנובא על סמך: (1) קוטר הפרח. (2) צפיפות קווי המתאר (אורך קו המתאר מחולק בשטח הפרח). (3) שטח הפרח, (4) שטח הפרח מחולק בקו המתאר. (5) המרחק שבין צבע הפרח לצבע הרקע במשושה הצבעים של דבורי בומבוס (טבלה 2).
לא נמצא מתאם (Pearson) מובהק בין מרחק הגילוי המירבי לבין: שטח הפרח, אורך קו המתאר, הניגודיות בירוק בין הפרח לרקע, ומרחק הצבע מהרקע (טבלה 2). גם רגרסיה לינארית מרובה שכללה את כל המשתנים הנ"ל לא הייתה מובהקת (F4,11=0.564, P=0.697), לכן ניתן לומר שמשתנים אלה, לבד או יחדיו, לא יכולים להסביר את מרחק הגילוי המירבי של הפרח.
מודל הרגרסיה המובהק היחיד לניבוי מרחק הגילוי המירבי של פרח על-ידי פועלות של דבורי בומבוס (איור 5) היה:
(MDD = A/(C*GC)
כאשר A= השטח הצבעוני של הפרח (סמ"ר), C=אורך קו המתאר של הפרח (ס"מ) ו GC=הניגודיות בין צבע הפרח לרקע באורך הגל הירוק.
טבלה 2. התוצאות של מבחני רגרסיה לינארית (ערך F, המובהקות P, והשונות המוסברת R2) לחיזוי של מרחק הגילוי המירבי של הפרחים (n=12) כמשתנה תלוי במשתנים הבלתי תלויים הבאים: קוטר הפרח, צפיפות קו המתאר, שטח, שטח חלקי קו המתאר, הניגודיות בירוק והמרחק מהמרכז במשושה הצבעים שחושבו כנגד הצבע הירוק כהה של המבוך. התוצאה המובהקת מוצגת באותיות מודגשות.
| R2 | P | F1,11 | משתנה תלוי |
| 0.1153 | 0.102 | 3.245 | קוטר (ס"מ) |
| 0.1238 | 0.264 | 1.403 | צפיפות קו המתאר (אורך קו המתאר חלקי שטח)(ס"מ/1) |
| 0.1228 | 0.264 | 1.399 | שטח (סמ"ר) |
| 0.2450 | 0.102 | 3.245 | שטח חלקי קו המתאר (ס"מ) |
| 0.0661 | 0.420 | 0.708 | הניגודיות בירוק בין צבע הפרח לצבע המבוך |
| 0.1523 | 0.210 | 1.797 | המרחק ממרכז משושה הצבעים המיצג את צבע המבוך |
| 0.3822 | 0.032 | 6.176 | שטח הפרח חלקי אורך קו המתאר והניגודיות בירוק (ס"מ) |
זווית הראייה המזערית (±ס.ת.) של 12 הפרחים שנבדקו הייתה 4.12±1.36 ס"מ (איור 4ב). לא נמצא מתאם מובהק בין זווית הראיה המזערית לבין מרחק הגילוי המירבי (Pearson r=-0.409, P=0.094). נמצא מתאם מובהק בין זווית הראיה המזערית לבין קוטר הפרח (Pearson r=0.605, P=0.0185) , ומתאם שלילי בינה לבין השטח של הפרח יחסית לשטח העיגול המקיף את דמות הפרח (Pearson r=-0.545, P=0.034). את זווית הראיה המזערית ניתן לנבא על פי משוואת הרגרסיה: זווית ראיה מזערית= 0.142*קוטר+ 1.371


דיון
במאמר זה אנו מציגים תוצאות ניסויים לקביעת המרחק המירבי ממנו פועלות של דבורי בומבוס יכולות לראות פרחים טבעיים, ואת זווית הראיה המזערית שלהן. נתונים אלה פורסמו לראשונה במאמר המקורי, המתורגם כאן. הקוטר הממוצע של הפרחים שנבדקו הוא כמעט שני ס"מ ומרחק הגילוי הממוצע הוא 28 ס"מ. כלומר, פועלות של דבורי בומבוס, שראייתן טובה מזו של דבורי דבש, חייבות להתקרב יותר מ 30 ס"מ על מנת לגלות פרח בודד שקוטרו כשני ס"מ. תוצאות אלה מתאימות לזה של המודל התיאורטי שהשתמש בצילומי פרחים וקבע שדבורים יכולות לגלות פרחים רק ממרחק קרוב מאוד (Vorobyev et al., 1997). זווית הראיה המזערית הממוצעת של הפרחים הייתה 4°, ולא היה קשר בינה לבין מרחק הגילוי המירבי. העדר הקשר הוא כנראה בשל העובדה שהמודלים התיאורטיים השתמשו במטרות נייר עגולות, ואילו הפרחים שנבדקו נבדלו זה מזה בצורתם ובמידת הניגודיות בצבע הירוק, למרות שהיו באותו קוטר. מכיוון שלמרחק הגילוי של פרחים השפעה על התנהגות שיחור המזון של דבורים בשדה ויעילותה, יש לו גם השפעה על הברירה הטבעית של תכונות הפרחים. לכן, ניתן לומר שמרחק הגילוי המרבי של פרחים טבעיים חשוב להבנת האקולוגיה והאבולוציה של פרחים ומאביקים, כמו שזווית הראיה המזערית חשובה למודלים הנוירופיזיולוגיים.
מרחק הגילוי של פרחים טבעיים בשדה הוא כנראה קצר עוד יותר מאשר זה שנמצא כאן בתנאי מעבדה. זאת בעיקר משום שבטבע הפרחים נגלים לדבורים מזוויות שונות, בהן נחשפים הפרחים כאשר שטחם קטן יותר מאשר לו היו נחשפים באופן חזיתי (Spaethe, 2001). בנוסף לכך הרקע הטבעי הוא רב-גוני ולכן סביר כי מקשה על גילוי של פרחים.
חוקרים רבים השתמשו במטרות נייר עגולות, והתחשבו בקוטר והצבע שלהן לקביעת זווית הראיה המזערית שלהן (Chittka & Raine, 2006; Giurfa et al., 1996; Giurfa & Vorobyev, 1997; Lehrer & Bischof, 1995; Menzel et al., 1997; Menzel & Giurfa, 2006; Vorobyev et al., 1997), וממנה חישבו את מרחק הגילוי המשוער, זאת למרות שמרבית הפרחים אינם עגולים אלא בעלי צורה גזורה ואז אין להם קוטר אחיד וברור. לכן, במחקר קודם (Ne’eman & Kevan, 2001) בדקנו את מרחק הגילוי של דגמים של פרחי נייר שנגזרו מנייר צבעוני כחול (לעיני אדם) שנבדלו בקוטר ובמידת הגזירות של צורתם על-ידי דבורי דבש. במחקר הקודם מצאנו שקוטר של מטרה דמוית פרח אינה יכולה להסביר את מרחק הגילוי שלה. מרחק הגילוי יכול היה להיות מוסבר ומנובא רק על פי השטח הצבעוני של הפרח בריבוע מחולק באורך קו המתאר של הפרח.
במחקר הנוכחי מצאנו שמרחק הגילוי של פרחים טבעיים על-ידי פועלות של דבורי בומבוס, אינו יכול להיות מנובא על-ידי אותה נוסחה (טבלה 2). יתכן שהסיבה לכך היא שהפרחים היו שונים במידת הניגודיות בצבע הירוק בינם לבין הרקע. זווית הראיה המזערית של מטרות עגולות על-ידי דבורי דבש עם ניגודיות בצבע הירוק היא 5º ושבע עיניות חייבות להיות מגורות על מנת שהדבורה תראה את המטרה. עבור מטרה נטולת ניגודיות כזו, הזווית היא 15º ו-59 עיניות חייבות להיות מגורות על מנת שהדבורה תראה את המטרה (Chittka & Raine, 2006; Giurfa & Vorobyev, 1997; Hempel de Ibarra et al., 2002). כלומר שמטרות בעלות ניגודיות בירוק נראות ממרחק גדול יותר מאשר מטרות דומות חסרות ניגודיות בירוק. עבור דבורי בומבוס זווית הראיה המזערית של מטרות עגולות עם ניגודיות בצבע הירוק היא 2.3º ושלוש עיניות חייבות להיות מגורות על מנת שהדבורה תראה את המטרה. זוית הראיה המזערית עבור מטרה נטולת ניגודיות כזו היא 2.7º ושבע עיניות חייבות להיות מגורות על מנת שהדבורה תראה את המטרה, והזויות משתנות גם בהתאם לגודל הגוף של הדבורה (Spaethe and Chittka, 2003). דבורי בומבוס מסוגלות לגלות פרחים ממרחק גדול יותר מאשר דבורי דבש, וההשפעה של הנגודיות בצבע הירוק קטנה יותר (Spaethe and Chittka, 2003; Dyer et al., 2008; Kapustjansky et al., 2010).
הניגודיות בצבע ירוק נמצאה חשובה בגילוי מטרות מרוחקות ואילו הניגודיות בכלל הצבעים בגילוי מטרות קרובות (Guirfa et al., 1996). הניגודיות בירוק נמצאה חשובה גם עבור ההפרדה של שלושה עיגולים סמוכים על-ידי דבורי דבש ודבורי בומבוס (Wertlen et al., 2008).
במחקר הנוכחי בדקנו את מרחק הגילוי של פרחים טבעיים על-ידי פועלות של דבורי בומבוס, ואנו מציעים משוואת רגרסיה לחיזוי מרחק הגילוי של פרחים על פי צורתם וצבעם. מודל הרגרסיה היחיד המאפשר חיזוי של מרחק הגילוי היה: MDD = A/(C*GC), כאשר A = השטח הצבעוני של הפרח (סמ"ר), C = אורך קו המתאר של הפרח (ס"מ) ו GC= הניגודיות בין צבע הפרח לרקע באורך הגל הירוק.
על מנת שדבורה תגלה מטרה, יש מספר מזערי של עיניות שחיבות להיות מגורות, וזה תלוי בגודל המטרה ובמרחק ממנה (Dyer et al., 2008; Giurfa and Vorobyev, 1997). לכן ניתן להניח, שלשטח הפרח השפעה ניכרת על מרחק הגילוי. אולם הגילוי שרק שטח הפרח בריבוע יכול להסביר את מרחק הגילוי אינו מתאים למודלים הנוכחיים של ראיית דבורים (Ne’eman & Kevan 2001). Bukovac et al., (2016). הראו שגם מרחק גילוי של עיגולי צבע במבוך דמוי Y אינו מוסבר טוב על-ידי המודלים הקיימים, אבל כן מתאים להסביר את האבולוציה של צבעי הפרחים. תגליות אלה מרמזות שגילוי פרחים מסובך יותר מאשר מציעים המודלים הנוירופיזיולוגיים הקיימים.
כאשר לשני פרחים שטח דומה אבל לאחד מהם קו מתאר ארוך יותר מאשר לשני, הוא גם בעל צורה גזורה יותר מאשר הפרח השני. כאשר שני פרחים הם בעלי קוטר דומה, אבל אחד בעל צורה גזורה יותר, הוא יגרה מספר קטן יותר של עיניות ולכן הדבורה חייבת להתקרב אליו יותר לפני שתוכל לראותו. זה ההסבר המוצע לעובדה כי אורך קו המתאר נמצא במכנה של משוואת הרגרסיה ומייצג יחס הפוך בינו לבין מרחק הגילוי.
ראיית צבע משמשת דבורים לראיה מטווח קרוב והניגודיות בירוק לראייה מרחוק. לכן, פרחים בעלי ניגודיות ירוקה אמורים להתגלות מוקדם יותר בזמן תעופה מאשר פרחים חסר ניגודיות בירוק (Spaethe & Chittka, 2003). הניגודיות בצבע ירוק של הפרחים במחקר זה היו בטווח בין 0.255 ל 0.499 (טבלה 1), כאשר הערך המירבי של ניגודיות הוא 0.5 (Spaethe et al. 2001). עובדה זו מסבירה את מיקומה של הניגודיות בירוק במכנה של משוואת הרגרסיה. העובדה שהניגודיות בירוק, (שהיא משתנה כמותי) הייתה דרושה על מנת לקבל משוואת רגרסיה מובהקת מרמזת על השפעה כמותית שלה. זאת בניגוד למודלים המציעים ערך סף שרק מעליו יש לניגודיות בירוק השפעה (Giurfa et al., 1996). במחקר אחר נמצאה קורלציה שלילית בין זמן החיפוש של פרחים קטנים מ 8 מ"מ לבין מידת הניגודיות בירוק (Spaethe et al. 2001). קורלציה שלילית נמצאה גם בין הניגודיות בירוק של פרחי דבורנית לבין משך הזמן של גילוי הפרח על-ידי זכרים של דבורים יחידאיות (Streinzer 2009), שתי דוגמאות אלה מדגימות גם הן השפעה כמותית של הניגודיות בירוק על התנהגות של דבורים. כמות סף של ניגודיות בירוק דרושה על מנת שדבורי דבש תהינה מסוגלות ללמוד שגמול מצוי בצורה גדולה, או קטנה, וליישם ידע זה גם לגבי צורות אחרות (Avarguès-Weber et al. 2014). למיטב ידיעתנו טרם נחקרה באופן ישיר ההשפעה הכמותית של הניגודיות בירוק על מרחק הגילוי של מטרות או פרחים.
משוואת הרגרסיה המוצעת לניבוי מרחק הגילוי של פרחים על-ידי דבורים מובהקת אבל מסבירה רק 38.2% מהשונות בנתוני מרחק הגילוי. כלומר, קיימים גורמים נוספים המשפיעים על מרחק הגילוי של פרחים שטרם נכללו במודל. אחד הגורמים האפשריים הוא גודל הדבורה. המבנה האופטי של העין בדבורת בומבוס נמצא בהתאמה לגודל גופה. גידול של 33% בגודל הגוף מעלה ב 100% את הרזולוציה של מטרה בודדת. מכיוון שדבורים גדולות הן בעלות עיניות גדולות יותר וראיה טובה יותר מאשר דבורים קטנות, יתכן שגודל גוף הדבורה הוא משתנה שיכול להעלות את אחוז השונות המוסברת במשוואת הרגרסיה.
תוצאות המחקר מראות כי ככל שפרחים הם בעלי צורה גזורה יותר. הם מתגלים לעין הדבורה רק ממרחק קצר יותר בהשוואה לפרחים מלאים , דבר העלול להוריד את תדירות ביקורי הדבורים וכתוצאה מכך להוריד את מידת הכשירות של הפרח. אם כך, מדוע לפרחים רבים צורה גזורה? דרכי צוף הם סימני צבע על עלי הכותרת של פרחים המנחים את המאביקים למיקום הגמול בפרח. דבורים מוצאות ושואבות את הצוף מהר יותר בפרחים בעלי דרכי צוף מאשר בפרחים חסרי דרכי צוף, וכתוצאה מכך ההסתברות להאבקה גדולה יותר בפרחים בעלי דרכי צוף מאשר בפרחים דומים חסרי דרכי צוף (Dafni & Kevan, 1997; Dafni et al., 1997; Dafni & Guirfa, 1999; Waser & Price, 1985). בנוסף ליכולת של דבורים מנוסות להשתמש בדרכי צוף, נמצא שדבורי בומבוס נאיביות, שלא ביקרו מעולם בפרח נמשכות לשולי הפרחים ועוקבות אחריהם, תוך שהם משתמשות בצורת הפרח כבדרכי צוף (Lunau et al., 2006). במחקר נוסף, נמצא שדבורי בומבוס מעדיפות לבקר בפרחים דמויי כוכב ובפרחים בעלי דרכי צוף רדיאליים, והעדפה זו נשארת גם לאחר הפסקת הגמול (Leonard & Papaj (2011. דרכי צוף מתגלות לדבורים רק לאחר הנחיתה בפרח, בעוד שפרחים דמויי כוכב מתגלים ממרחק גדול יותר. לכן הועלתה ההצעה שפרחים גזורים דמויי כוכב עשויים להנחות דבורים לגמול בפרח בדומה לדרכי צוף (Dafni & Kevan, 1997). אנליזה של כל פרחי ישראל הראתה שלפרחים קטנים (< 5cm2) קו מתאר ארוך יותר ומראה גזור יותר מאשר לפרחים גדולים (Menzel & Shmida 1993). לכן אנו מציעים שפרחים קטנים מפצים בצורתם, הגזורה באופן רדיאלי, על מרחק הגילוי הקצר שלהם על-ידי דבורים, ושלפרחים גזורים יהיו פחות דרכי צוף מאשר לפרחים רדיאליים בלתי גזורים.
לסיכום, אנו מציעים כאן נוסחה ניסיונית היכולה לשמש לחיזוי מרחק הגילוי של פרחים על-ידי פועלות של דבורי בומבוס, בעזרת פרמטרים של צורת הפרח והצבע שלו. מודל זה עדיין דורש שיפורים ועידון שיושגו על-ידי שימוש בטיפוסי פרחים נוספים ושונים, ובתנאים המדמים טוב יותר את הסביבה הטבעית. פיתוח שיטות לבדיקת מרחק הגילוי של פרחים בשדה הוא מסובך ונותר אתגר עתידי. קביעה ניסויית של מרחק הגילוי המזערי היא חשובה, וטוב אם יעשו בו שימוש במחקרים עתידיים על התנהגות שיחור המזון של דבורים בסביבה טבעית, ובמקום בו פורחים פרחים שונים בו זמנית. מרחק הגילוי של פרחים חשוב להבנה של השפעת המאביקים על האבולוציה של הצורה והצבע של פרחים, ולצורך בדיקתם של מודלים נוירופיסיולוגיים המנסים להסביר את המנגנונים של ראיית צורה וצבע של פרחים.
תודות
תודתנו נתונה ל Lars Chittka, על האירוח במעבדתו ב Queen Mary University of London, על הצעותיו, הערותיו ותמיכתו בביצוע המחקר. תודה גם ל A. G. Dyer על הערותיו.
ספרות:
Anderson AM 1977 Shape perception in the honey bee. Animal Behaviour 25: 67-79.
Armbruster WS Antonsen L and Pélabon C 2005 Phenotypic selection on Dalechampia blossoms: honest signalling affects pollination success. Ecology 86: 3323-3333.
Avarguès-Weber A Deisig N and Giurfa M 2011 Visual cognition in social insects. Annual Review of Entomology 56: 423-443.
Avarguès-Weber A and Giurfa M 2014 Cognitive components of color vision in honey bees: how conditioning variables modulate color learning and discrimination. Journal of Comparative Physiology A 200: 449-461.
Avarguès-Weber A d’Amaro D Metzler M and Dyer AG 2014 Conceptualization of relative size by honeybees. Frontiers in Behavioral Neurosciences 8: 80.
Backhaus W 1991 Color opponent coding in the visual system of the honeybee. Vision research 31: 1381–1397.
Blarer A Keasar T and Shmida A 2002 Possible mechanisms for the formation of flower size preferences by foraging bumblebees. Ethology 108: 341-351.
Bukovac Z Dorin A and Dyer A 2013 A-Bees See: a simulation to assess social bee visual attention during complex search tasks. In: Lio P Miglino O Nicosia G Nolfi S Pavone M (eds) Advances in artificial life, ECAL 2013. Proceedings of the twelfth European conference on the synthesis and simulation of living systems, Taormina, September 2013. Complex Adaptive Systems, MIT Press, Cambridge, London, pp 276–283.
Bukovac Z Dorin A Finke V Shrestha M et al. 2016 Assessing the ecological significance of bee visual detection and colour discrimination on the evolution of flower colours. Evolutionary Ecology 30: 1-20.
Chittka L 1992 The colour hexagon: a chromaticity diagram based on photoreceptor excitations as a generalized representation of colour opponency. Journal of Comparative Physiology A 170: 533-543.
Chittka L and Raine NE 2006 Recognition of flowers by pollinators. Current Opinion in Plant Biology 9: 428-435.
Chittka L and Spaethe J 2007 Visual search and the importance of time in complex decision making by bees. Arthropod-Plant Interactions 1: 37-44.
Conner JK and Rush S 1996 Effects of flower size and number on pollinator visitation to wild radish, Raphanus raphanistrum. Oecologia 105: 509-516.
Dafni A and Giurfa M 1999 The functional ecology of floral guides in relation to insects behaviour and vision. In: Waser S (ed) Evolutionary theory and processes: modern perspectives. Kluwer Academic Publishers, Dordrecht, pp. 363-383.
Dafni A and Kevan PG 1997 Flower size and shape: implications in pollination. Israel Journal of Plant Sciences 45: 201-211.
Dafni A Lehrer M and Kevan P 1997 Spatial flower parameters and insect spatial vision. Biological Review 72: 239-282.
Dyer AG and Chittka L 2004 Biological significance of distinguishing between similarcolours in spectrally variable illumination: bumblebees (Bombus terrestris) as a case study. Journal of Comparative Physiology A 190: 105-114.
Dyer AG Whitney HM Arnold SE Glover BJ et al. 2007 Mutations perturbing petal cell shape and anthocyanin synthesis influence bumblebee perception of Antirrhinum majus flower colour. Arthropod-Plant Interactions 1: 45-55.
Dyer AG Spaethe J and Prack S 2008 Comparative psychophysics of bumblebee and honeybee colour discrimination and object detection. Journal of Comparative Physiology A 194: 617-627.
Dyer AG Streinzer M and Garcia J 2016 Flower detection and acuity of the Australian native stingless bee Tetragonula carbonaria Sm. Journal of Comparative Physiology A, 202: 629-639.
Elle E and Carney R 2003 Reproductive assurance varies with flower size in Collinsia parviflora (Scrophulariaceae). American Journal of Botany 90: 888-896.
Fenster CB Cheely G Dudash MR and Reynolds RJ 2006 Nectar reward and advertisement in hummingbird-pollinated Silene virginica (Caryophyllaceae). American Journal of Botany 93: 1800-1807.
Frisch Kv 1914 Der Farbensinn und Formensinn der Biene. Gustav Fischer, Jena.
Galen C 1999 Why do flowers vary? The functional ecology of variation in flower size and form within natural plant populations. Bioscience 49: 631-640.
Galen C and Newport M 1987 Bumble bee behavior and selection on flower size in the sky pilot, Polemoniumviscosum. Oecologia 74: 20-23.
Giurfa M and Vorobyev M 1997 The detection and recognition of color stimuli by honeybees: performance and mechanisms. Israel Journal of Plant Sciences 45:129-140.
Giurfa M Vorobyev M Kevan P and Menzel R 1996 Detection of coloured stimuli by honeybees: minimum visual angles and receptor specific contrasts. Journal of Comparative Physiology A 178: 699-709.
Goodwillie C Sargent RD Eckert CG Elle E et al. 2010 Correlated evolution of mating system and floral display traits in flowering plants and its implications for the distribution of mating system variation. New Phytologist. 185: 311-321.
Heinrich B 1975 Energetics of pollination. Annual Review of Ecology and Systematics 15: 139-170.
Hempel de Ibarra NH Giurfa M and Vorobyev M 2001 Detection of coloured patterns by honeybees through chromatic and achromatic cues. Journal of Comparative Physiology A 187: 215-224.
Hempel de Ibarra NH Giurfa M and Vorobyev M 2002 Discrimination of coloured patterns by honeybees through chromatic and achromatic cues. Journal of Comparative Physiology A 188: 503-512.
Hempel de Ibarra NH Vorobyev M and Menzel R 2014 Mechanisms, functions and ecology of colour vision in the honeybee. Journal of Comparative Physiology A 200: 411-433.
Hempel de Ibarra NH Langridge KV and Vorobyev M 2015 More than colour attraction: behavioural functions of flower patterns. Current Opinion in Insect Science 12: 64-70.
Herrera J 2005 Flower size variation in Rosmarinus officinalis: individuals, populations and habitats. Annals of Botany 95: 431-437.
Hertz M 1929 Die Organisation des optischen Feldes bei der Biene. Journal of Comparative Physiology A 8: 693-748.
IBM Corporation 2010 IBM SPSS Statistics for Windows, Version 19.0. Armonk, NY IBM Corp.
Kaczorowski RL Seliger AR Gaskett AC Wigsten SK et al. 2012 Corolla shape vs. size in flower choice by a nocturnal hawkmoth pollinator. Functional Ecology 26: 577-587.
Kapustjansky A Chittka L and Spaethe J 2010 Bees use three-dimensional information to improve target detection. Naturwissenschaften 97: 229-233.
Keasar T Pollak G Arnon R Cohen D and Shmida A 2006 Honesty of signalling and pollinator attraction: the case of flag-like bracts. Israel Journal of Plant Sciences 54: 119-128.
Krizek BA and Anderson JT 2013 Control of flower size. Journal of Experimental Botany 64: 1427-1437.
Kugler H 1934 Blütenokologische Untersuchungen mit Hummeln VII Die Anlockung von "Neulingen" durch Blüten. Planta 23: 692-714.
Lehrer M and Bischof S 1995 Detection of model flowers by honeybees: the role of chromatic and achromatic contrast. Naturwissenschaften 82: 145-147.
Lehrer M Horridge GA Zhang SW and Gadagkar R 1995 Shape vision in bees: Innate preference for flower-like patterns. Philosophical Transactions of the Royal Society B Biological Sciences 347: 123–137.
Leonard AS Papaj DR 2011 ‘X’ marks the spot: the possible benefits of nectar guides to bees and plants. Functional Ecology 25: 1293-1301.
Lunau K Fieselmann G Heuschen B and van de Loo A 2006 Visual targeting of components of floral colour patterns in flower-naive bumblebees (Bombus terrestris; Apidae). Naturwissenschaften 93: 325-328.
Martin NH 2004 Flower size preferences of the honeybee (Apis mellifera) foraging on Mimulus guttatus (Scrophulariaceae). Evolutionary Ecological Research 6: 777-782.
Menzel R and Giurfa M 2006 Dimensions of cognition in an insect, the honeybee. Behavioural and Cognitive Neuroscience Reviews 5: 24-40.
Menzel R Gumbert A Kunze J Shmida A et al. 1997 Pollinators' strategies in finding flowers. Israel Journal of Plant Sciences 45: 141-156.
Menzel R and Shmida A 1993 The ecology of flower colours and the natural colour vision of insect pollinators: the Israeli flora as a study case. Biological Review 68: 81-120.
Neal PR Dafni A and Giurfa M 1998 Floral symmetry and its role in plant-pollinator systems: terminology, distribution, and hypotheses. Annual Review of Ecology and Systematics 29: 345-373.
Ne’eman G and Kevan PG 2001 The effect of shape parameters on maximal detection distance of model target by honeybee workers. Journal of Comparative Physiology A 187: 653-660.
Peitsch D Fietz A Hertel H de Souza J et al. 1992 The spectral input systems of hymenopteran insects and their receptor-based colour vision. Journal of Comparative Physiology A 170: 23–40.
Plowright CMS Evans SA Leung JC and Collin CA 2011 The preference for symmetry in flower-naïve and not-so-naïve bumblebees. Learning and Motivation 42: 76-83.
Schaefer HM Schaefer V and Levey DJ 2004 How plant–animal interactions signal new insights in communication. Trends in Ecology and Evolution 19: 577-584.
Simonds V and Plowright CMS 2004 How do bumblebees first find flowers? Unlearned approach responses and habituation. Animal Behaviour 67: 379-386.
Spaethe J and Chittka L 2003 Interindividual variation of eye optics and single object resolution in bumblebees. Journal of Experimental Biology 206: 3447-3453.
Spaethe J Tautz J and Chittka L 2001 Visual constraints in foraging bumblebees: flower size and color affect search time and flight behavior. Proceedings of The National Academy of Scieinces Biology 98: 3898-3903.
Sprengel CK 1793/1996 Discovery of the secret of nature in the structure and fertilization of flowers (translated by Hasse P) in: Lloyd D G Barrett S C (eds.) Floral biology: studies on floral evolution in animal-pollinated plants. Springer, New York, pp 3-43..
Srinivasan MV 2010 Honey bees as a model for vision, perception, and cognition. Annual Review of Entomology 55: 267-284.
Streinzer M Paulus HF and Spaethe J 2009 Floral colour signal increases short-range detectability of a sexually deceptive orchid to its bee pollinator. Journal of Experimental Biology 212: 1365-1370.
Thompson EL and Plowright CM 2014 How images may or may not represent flowers: picture–object correspondence in bumblebees (Bombus impatiens)? Animal Cognition 17: 1031-1043.
Vorobyev M Gumbert A Kunze J Giurfa M et al. 1997 Flowers through insect eyes. Israel Journal of Plant Sciences 45: 93-101.
Waser NM and Price MV 1985 The effect of nectar guides on pollinator preference: experimental studies with a montane herb. Oecologia 67: 121-126.
Wertlen AM Niggebrugge C Vorobyev M Hempel and de Ibarra N 2008 Detection of patches of coloured discs by bees. Journal of Experimental Biology 211: 2101-2104.
Wu W Moreno AM Tangen JM and Reinhard J 2013 Honeybees can discriminate between Monet and Picasso paintings. Journal of Comparative Physiology A 199:45-55.
Zhang S Srinivasan MV Zhu H and Wong J 2004 Grouping of visual objects by honeybees. Journal of Experimental Biology 207: 3289-3298.
=====================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: נאמן ג ונאמן ר 2017 מאיזה מרחק רואה דבורה פרח? מרחק הגילוי המירבי של פרחים טבעיים על-ידי דבורי בומבוס, כתב-עת "כלנית" מספר 4.









