

גידי נאמן – החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים gneeman@gmail.com
תקציר: במקומות עם אקלים ים-תיכוני, האביב הוא עונת השיא של הפריחה ושל שפע המאביקים. למרות זאת, יש מספר מיני גאופיטים הפורחים בסתיו, טרם תחילת הגשמים שזו התקופה היבשה ביותר בשנה. הסבר מקובל לפריחה יוצאת דופן זו, הוא שמינים אלה התפתחו ממינים פורחי-אביב שהסיטו את תקופת הפריחה שלהם לסתיו. על-פי סברה זו, הגורם הסלקטיבי היכול להניע שינוי כזה, הוא הצלחה רבייתית גדולה יותר בסתיו. זאת בשל מיעוט מיני הצמחים הפורחים, והתחרות הנמוכה על מאביקים, לעומת התחרות הגבוהה אביב. מטרת מחקר זה היא לבחון את ההשערה שההסטה של מועד הפריחה מהאביב לסתיו בחצב מצוי Urginea mritima הייתה מלווה בעליה בהצלחה הרבייתית. מכיוון שאין חצבים הפורחים באביב, נאלצנו להשוות בין החצב המצוי לבין בן-חצב יקינטוני Scila hyacintoides שהוא מין קרוב הדומה לחצב בתכונות רבות, אך פורח באביב. ההשוואה כללה בדיקה של ההצלחה הרבייתית הנקבית והזכרית, מספר גרגירי האבקה והביציות המיוצרים בפרח, מנגנון הרבייה וההאבקה, וההשקעה בביומסה באיברים הווגטטיביים, בפרחים למשיכת מאביקים ובזרעים. התוצאות מלמדות על יתרון יחסי בהצלחה הזכרית של חצב מצוי יחסית לזו של בן-חצב יקינטוני. מבחינה זו התוצאות תומכות בהשערה שהסטת הפריחה מהאביב לסתיו הייתה מלווה ביתרון רבייתי. אבל להסטה זו היה מחיר בהגדלת ההשקעה בפרחים וירידה בהצלחה הנקבית בחצב מצוי יחסית לבן חצב יקינטוני. אולם יתכן שהיתרון בפריחה הסתווית טמון בטריפת זרעים נמוכה יותר והצלחה גדולה יותר בנביטה והתבססות הצמחים, נושאים אלה מוצעים למחקרים בעתיד.
מבוסס על המאמר:
Ne’eman G 1999 A comparative study of reproduction and biomass allocation between spring and autumn flowering Mediterranean geophytes. Journal of Mediterranean Ecology 1:27-34.
מבוא
גאופיטים, או בשמם המקורי קריפטופיטים (Raunkiaer 1934), הם צמחים המסוגלים לעבור תקופות קשות של יובש או קור בעזרת ניצני התחדשות הנמצאים על אברים אוגרי מזון תת-קרקעיים כמו פקעות, בצלים, או קנה שורש (Archibold 1995). צמחים בעלי צורת-חיים זו נמצאים במקומות שונים בעולם, בתצורות צומח שונות. בחלקים היובשניים של החבל הים-תיכוני בישראל הם מהווים 8-17% ממיני הצמחים (Orshan 1983, Shmida & Dafni 1989). הגאופיטים בישראל מלבלבים לאחר רדת גשמי הסתיו, בחורף ובאביב העלים צומחים, מטמיעים ומייצרים מזון הנאגר באברים התת-קרקעיים, זאת זמן שיש מים זמינים בקרקע ותנאי הסביבה נוחים (גליל 1961). העלים מתייבשים בתחילת הקיץ היבש (Dafni et al. 1981a), ולכן הם בעצם עומדים בשלכת קיץ.
גאופיטים שונים פורחים בישראל כמעט במשך כל השנה. מרבית המינים פורחים בשיא עונת הפריחה באביב, חלקם פורחים בסוף הקיץ ובסתיו, ואחרים במשך החורף המתון (Shmida & Dafni 1989). מרבית מיני הגאופיטים פורחים לאחר הגשמים כשהעלים מלווים את הפרחים, לכן הם מכונים סינאנתיים (synanthous) שפרושו במקביל לפרחים (Dafni et al. 1981a). בניגוד למינים אלה, יש קבוצה של גאופיטים הפורחים בסוף הקיץ ובסתיו לפני רדת היורה בתקופה היבשה ביותר בשנה. גאופיטים אלה מהווים כ 80% מכלל מיני הצמחים פורחי-הסתיו בישראל (Shmida & Dafni 1989). הגאופיטים בקבוצה זו פורחים ללא עלים המופיעים מיד לאחר תחילת הגשמים, ונקראים היסטראנתיים (hysteranthous) (Dafni et al. 1981a). המינים בקבוצה זו, שפרחיהם נישאים על עמוד פריחה ואינם צמודי-קרקע, מפזרים את זרעיהם בסתיו ואלה נובטים מיד לאחר רדת הגשמים (Dafni et al. 1981b).
האביב הוא שיא עונת הפריחה של הצמחים מכל צורות החיים (עשבוניים, שיחים ועצים) בארצות הים התיכון. זאת, בשל ההשפעה של הקיץ החם הארוך והיבש על מחזור חיי הצמחים, ובשל שפע המאביקים בעונה זו (Zohary 1976, Auerbuch & Shmida 1987, Shmida & Dafni 1989). אבל, למרות שהמגוון והשפע של המאביקים הוא הגדול ביותר באביב, עדיין יתכן שיש מחסור יחסי של מאביקים לכל פרח, בשל שפע הפריחה. לכן המינים השונים מתחרים על שירותי המאביקים על-ידי הגדלת הפרסומת של פרחיהם והגמול הניתן בהם למאביקיהם (Cohen & Smimida 1993).
התחרות על שירותי מאביקים מהווה גורם סלקטיבי המשפיע על האבולוציה והקביעה של עונת הפריחה של מיני צמחים שונים באופן המצמצם את התחרות, ומגדיל את הכשירות של צמח הפורח בעונה בה התחרות נמוכה יותר (Mosquin 1971, Heinrich & Raven 1972). השערה זו נתמכת על-ידי תוצאות של ניסויים המדגימים שתחרות גבוהה על שרותי המאביקים גורמת לירידה בתדירות ביקורי המאביקים וירידה בשיעור העברת אבקה קונספציפית, כלומר בין פרחים השייכים לאותו מין (Gentry 1964, Waser 1979, 1983, Pleasant 1980, Campbell & Motten1985). אבל נראה, שתחרות על מאביקים אינה יכולה להסיט את מועד הפריחה מעבר לגבולות הנקבעים על-ידי גורמי סביבה ואילוצים פילוגנטיים (Kochmer & Handel 1986).
בדרום אפריקה באזור המעבר בין החבל האקלימי הים-תיכוני לבין החבל הטרופי-יובשני, נמצאות אוכלוסיות צמחים הנבדלות במועדי הפריחה שלהן (Letty et al. 1962), עדות לכך שמועד הפריחה יכול להשתנות כתוצאה מהברירה הטבעית. בנוסף לכך נמצא גם שמועד הפריחה של צמחי-בר וצמחי-תרבות יכולים להשתנות כשהם גדלים בתנאים השונים מאלה שבתחום תפוצתם הטבעי (waser 1983).
דפני וחבריו (Dafni et al. 1981b) הציעו שצמחים היסטראנתיים, הפורחים ללא עלים, התפתחו אבולוציונית מצמחים סינאנתיים, הפורחים עם עלים. בבני קבוצת החצב (הפורחים עם עמוד פריחה) נדחה מועד הפריחה מהאביב, ובמינים בני קבוצת הכרכום (שפרחיהם צמודי-קרקע) הוקדם מועד הפריחה. פריחה ללא עלים התפתחה במקביל במספר סוגי צמחים כמו רקפת, בן-חצב וחצב, שכל אחד מהם מכיל גם מינים הפורחים עם עלים ומינים הפורחים ללא עלים (Dafni et al. 1981b).
מרבית האוכלוסיות של הרקפת המצויה והנרקיס המצוי בישראל, פורחים בחורף לאחר שהצמיחו עלים. אבל, בבתי גידול סלעיים ויבשים נמצאות אוכלוסיות הפורחות בסתיו ללא עלים (Arroyo & Dafni 1995, Schwartz-Tzachor et al. 2008).
השקעה ברבייה, צמיחה ואחזקה, הם שלושה תחומים חליפיים הקובעים את אסטרטגיות החיים, צורת החיים וההתנהגות העונתית של צמחים שונים (Willson 1983). בהתאם לתאוריה של השקעת אנרגיה אופטימלית (optimal allocation theory) (Enquist et al. 1999), צמחים מטמיעים כשהתנאים לכך מיטביים, הם אוגרים חומרים עתירי אנרגיה ומשתמשים בה בזמן המתאים (Bloom et al. 1985, Kozlowski 1992). ההשקעה ברבייה כרוכה בהשקעה ניכרת של אנרגיה, לכן צמחים רבים אוגרים חומרי מזון עתירי אנרגיה, ומשתמשים בהם לייצור פרחים ורבייה במועד האופטימלי מבחינת תנאי סביבה והסיכויים להאבקה (Heinrich 1975). ההשקעה ברבייה עשויה להיות על חשבון אנרגיה אגורה, צמיחה בהווה ובעתיד, ולעיתים אפילו על חשבון הישרדות (Shon & Policanski 1977, Evenson 1983, Primack & Stacy 1998).
כאמור, הצמחים פורחי-הסתיו התפתחו אבולוציונית כנראה מצמחים פורחי-אביב. מכיוון שתנאי הסביבה בסוף הקיץ ובסתיו הם קשים ביותר מבחינת חום ויובש, התפתחות אבולוציונית כזו, סותרת לכאורה את תאורית ההשקעה האופטימלית. זאת, אלא אם מעבר כזה היה כרוך ברווח בתחום אקולוגי אחר, אשר העלה את הכשירות של הצמחים, והקנה להם יתרון תחרותי על פני צמחים אחרים פורחי-אביב. שלושה גורמים כאלו הם: 1. תחרות נמוכה יותר על מאביקים בסתיו, הגורמת לעליה בפוטנציאל הרבייה של הצמחים. 2. ירידה בהעברת אבקה בין צמחים בני מינים שונים הפוגעת בפוטנציאל הרבייה של הצמח, זאת בשל נאמנות הדבורים והמספר הנמוך של המינים הפורחים בסתיו (Dafni et al. 1981b, Shmida & Dafni 1989). 3. הפחתה בטריפת הזרעים המפוזרים בסתיו ונובטים מיד לאחר רדת הגשמים, יחסית לזרעים המפוזרים באביב וחשופים לטריפה למעלה מחצי שנה. בשל העובדה שלכל מין עונת פריחה משלו, ההשוואה בין פורחי-אביב לפורחי-סתיו קשה וחסרים מחקרי שדה המראים יתרון בכשירות של צמחים פורחי-סתיו על פני פורחי-אביב.
בהעדר אפשרות השוואה של אוכלוסיות שונות בנות אותו מין, מה שנותר הוא לשוות בין מינים קרובים מבחינה טקסונומית, הדומים גם באסטרטגיית החיים שלהם, כשאחד פורח באביב והשני בסתיו. מטרת מחקר זה היא לבדוק את ההשערה שהמעבר מפריחת אביב לפריחת סתיו הקנה יתרון רבייתי לחצב מצוי הפורח בסתיו לעומת קרובו בן-חצב יקינטוני הפורח באביב.
חומרים ושיטות
צמחי המחקר
בן חצב יקינטוני (Scilla hyacantoid,es L.) (איור 1) ממשפחת האספרגיים (לשעבר השושניים), הוא גאופיט הגדל ביערות וחורשים פתוחים של אלון מצוי ואלון תבור בצפונה של ישראל. לצמח בוגר יש בצל תת-קרקעי והוא מפתח, מיד לאחר תחילת גשמי הסתיו, שושנת עלים בת 10 עלים, כל אחד ברוחב של כ 5 ס"מ ואורך של כ 30 ס"מ. עמוד התפרחת מתחיל לצמוח בסוף פברואר ומגיע לגובה של כמטר אחד, והוא נושא כ- 150פרחים פתוחים בצבע תכלת ובקוטר של כס"מ אחד. הפרחים מסודרים בדורים, ונפתחים מתחתית העמוד למעלה במשך כל זמן הפריחה בחודשים אפריל ומאי. הפרחים מבוקרים על-ידי דבורי דבש, דבורים יחידאיות קטנות ועל-ידי זבובים. בתחילת הקיץ, לאחר הבשלת הזרעים ופיזורם, מתייבשים עמוד התפרחת והעלים והצמח נמצא בתרדמה ושלכת קיץ עד לגשמי הסתיו.


איור 1. בן-חצב יקינטוני תמונה כללית ותקריב. צילום: עוזי פז ©
חצב מצוי (Urginea maritima (L.) BAKER, syn. Drimia maritima (L.) Stearn) (איור 2) ממשפחת האספרגיים (לשעבר השושניים) (Plants of the world online 2020), הוא גאופיט נפוץ מאוד בשטחים עשבוניים ושטחי בתה פתוחים כולל שטחים מופרים. בישראל הוא גדל מהגליל העליון בצפון לאורך מפל הגשם, ועד לבתי גידול סלעיים במפנים צפוניים במדבר יהודה והר הנגב. לצמחים בוגרים בצל רדוד המגיע עד לקוטר של כ- 20 ס"מ. מיד לאחר תחילת הגשמים, מצמיח הצמח שושנת עלים בת כ 10 עלים , כל אחד ברוחב של כ- 10 ס"מ ובאורך של כ- 50 ס"מ. העלים ירוקים ומטמיעים במשך כל החורף והאביב עד לתחילת היובש בקיץ. כבר בסוף הקיץ החל מסוף אוגוסט, הצמח מתחיל לגדל עמוד תפרחת המגיע עד לגובה של כמטר וחצי, הוא נושא עד כ -300 פרחים לבנים פתוחים בקוטר של כס"מ אחד. הפרחים מבוקרים על-ידי דבורי דבש, צרעות, וזבובים, ומאבקים גם על-ידי הרוח (Dafni & Dukas 1986). הפירות מבשילים במהירות והזרעים מופצים עד סוף אוקטובר. הזרעים חסרי תרדמה ונובטים מיד לאחר גשמי הסתיו.


איור 2. חצב מצוי תמונה כללית ותקריב. צילום: עוזי פז ©
ניטור באתרי המחקר
המחקר נערך בגליל העליון והתחתון. התצפיות בבן-חצב יקינטוני נערכו באמירים ברום של כ 500 מ' וליד אלון הגליל ברום של כ 200 מ', והתצפיות בחצב מצוי נערכו ליד שורשים ברום של כ 200 מ' וליד טבעון ברום של כ 100 מ'. בכל אתר נערכו תצפיות ב 30-40 צמחים מסומנים שהיו חשופים להאבקה טבעית, החל מאוגוסט 1990 עד יוני 1991 . הניטור כלל: 1. מדידה לא הרסנית של קוטר הבצל (על-ידי חשיפת חציו העליון של הבצל). 2. ספירת העלים. 3. מדידת שטח העלים הבוגרים על-ידי העתקת צורתם לנייר, גזירת הנייר, שקילת הנייר וחלוקה במשקל של סמ"ר אחד של נייר. 4. מדידת הגובה והקוטר של עמוד התפרחת. 5. ספירת הפרחים (Fl). 6. ספירת הפירות (Fr). 7. ספירת מספר הזרעים (S) ב 10 הלקטים שנבחרו באופן אקראי, וחישוב מספר הזרעים הממוצע לפרי. 8. חישוב אחוז חנטת הפירות בכל צמח והממוצע בכל אוכלוסייה.
האבקה, חנטת פירות ויצור זרעים
בכל מין נבחרו באקראי וסומנו 20 צמחים. 40 פרחים בכל צמח, לא טופלו ונבדקו כביקורת להאבקה חופשית. 40 פרחים אחרים על אותו צמח שהיו חשופים לביקורי מאביקים, הואבקו ידנית באבקה זרה בבוקר של היום הראשון לפריחתם, כטיפול של האבקה משלימה. 40 פרחים בחמישה צמחים מכל מין כוסו ברשת עדינה (גודל עין 1מ"מ) לפני פתיחת הפרחים למניעת ביקורי חרקים, לבדיקת קיום של האבקה עצמית ספונטנית או יצירת זרעים ללא הפרייה. 40 פרחים נוספים על אותם צמחים כוסו באותה ברשת לפני פתיחת הפרחים, פרחים אלה הואבקו ידנית באבקה זרה ביום הראשון לפריחתם, לבדיקת הפוטנציאל המכסימלי של הרבייה והאפקטיביות של טיפול ההאבקה המשלימה. אחוז חנטת הפירות ומספר הזרעים בהלקט, שימשו כמדד להשוואת ההשפעה של הטיפולים השונים.
אבקה, ביציות, זרעים והצלחה רבייתית
מכל מין נאספו עשרים כפתורי פרחים טרם פתיחתם ושומרו ב FAA (חומר משמר), לבדיקה במעבדה. השחלות של כל הפרחים נבדקו תחת בינוקולר ונספרו הביציות (O) בכל שחלה. שני מאבקים מכל פרח פורקו וגרגירי האבקה נצבעו והורחפו בנפח מדוד של נוזל. גרגרי האבקה נספרו בדגימה בנפח ידוע תחת מיקרוסקופ בהמוציטומטר (המשמש לספירת כדוריות דם) וחושב מספר גרגרי האבקה (P) בכל פרח שלו שישה אבקנים. יחס O/P (מספר גרגירי האבקה יחסית למספר הביציות) חושב לכל פרח וחושב הממוצע לכל מין. היחס בין מספר הזרעים לבין מספר הביציות חושב לכל מין. ההצלחה הרבייתית היחסית (RRS relative reproductive success) חושבה בנוסחה RRS=(Fr/Fl)*S/O (Wiens 1984),כאשר Fr הוא מספר הפירות, Fl הוא מספר הפרחים, S מספר הזרעים ו O מספר הביציות.
השקעת ביומסה
עשרה בצלים מכל מין נאספו בחורף, הקוטר שלהם נמדד לפני יבושם במשך כשבוע בטמפרטורה של 80ºC עד להגעתם למשקל יבש יציב לקביעת הביומסה שלהם. נוסחת הרגרסיה של הביומסה כנגד הקוטר, חושבה להערכת הביומסה של הבצלים החיים של הצמחים המנוטרים שקוטרם נמדד בשדה. באופן דומה יובשו 10 סמ"ר של עלה מכל מין, וחושבה הביומסה של העלים של הצמחים המנוטרים בשדה על פי שטחם. באופן דומה נקבעה גם הביומסה של עמוד התפרחת של הצמחים המנוטרים, לאחר יבוש וקביעת הביומסה של 10 ס"מ של גבעול פריחה. יובשו פרחים וזרעים ונקבעה הביומסה הממוצעת של פרח וזרע בודד של כל מין. נתונים אלה שימשו לחישוב ההשקעה בביומסה האגורה בבצלים, זו המושקעת בעלים וזו המושקעת בפריחה ובזרעים.
ניתוח הנתונים
השפעת הטיפולים בתוך כל מין נבדקה על-ידי ניתוח שונויות חד-כיווני (One way ANOVA). במקרה של השפעה מבהקת (P<0.05) בוצע מבחן Duncan’s multiple range test להשוואת ההשפעה בין הטיפולים השונים. מבחן T בוצע להשוואה של כל משתנה בין שני המינים. מקדם השונות CV חושב (CV=STD/mean) להשוואת מידת השונות של משתנים בין המינים. במקרים בהם לא התגלתה השפעה מובהקת של הטיפול על משתנה הנמדד בשני המינים, אוחדו הנתונים של הטיפול והביקורת בכל מין ולאחר מכן מערך מבחן T להשוואה בין שני המינים. כל המבחנים בוצעו לאחר טרנספורמציה arcsin של הפרופורציות במבחן GLM ומבחן T בתכנת SAS (SAS 1988).
תוצאות
האבקה, חנטת פירות ויצור זרעים
חנטת הפירות (טבלה 1) (ממוצע ± סטיית תקן) הייתה 89%±11.4% בבן-חצב יקינטוני, ו 35.5%±30.0 בחצב מצוי, והבדל זה היה מובהק (t(55.1)=10.1, P=0.0001). הערכים הממוצעים של כל שאר המשתנים שנבדקו בשני המינים מוצגים בטבלה 1.
טבלה 1. המספר הממוצע (± סטיית תקן) של גרגירי האבקה לפרח, הביציות לפרח, הזרעים להלקט (פרי) היחס בין מספר גרגרי האבקה למספר הביציות לפרח (O/P), היחס בין מספר הפרחים לפירות לצמח (Fr/Fl), היחס בין מספר הזרעים לביציות (*S/O) וההצלחה הרבייתית היחסית לצמח (RRS*) . *הערך מחושב מהמוצעים.
| בן-חצב יקינטוני | חצב מצוי | |||
| המשתנה | ממוצע (± ס.ת.) | N | ממוצע (± ס.ת.) | n |
| גרגרי אבקה (P) | 21,060±2,652 | 40 | 26,580±2,977 | 20 |
| ביציות (O) | 6.3±1.2 | 40 | 39.6±2.6 | 20 |
| זרעים (S) | 3.83±0.97 | 67 | 9.33±3.58 | 68 |
| P/O | 3,435±618 | 40 | 676±100 | 20 |
| Fr/Fl | 0.890±0.114 | 70 | 0.355±0.300 | 80 |
| *S/O | 0.513 | 0.221 | ||
| RRS* | 0.4566 | 0.0785 |
בחצב מצוי לא נמצאו הבדלים מובהקים בחנטת הפירות בין טיפולי ההאבקה השונים אבל בבן-חצב יקינטוני ההבדלים היו מובהקים. חנטת הפירות כתוצאה מהאבקה עצמית ספונטנית, או רוח הייתה נמוכה מאוד בשני המינים, והאבקה משלימה לא העלתה את חנטת הפירות יחסית להאבקה פתוחה (איור 3). מקדם השונות בחנטת הפירות של חצב מצוי (0.845=CV) היה גבוה בהרבה מזה של בן חצב יקינטוני (0.128=CV).
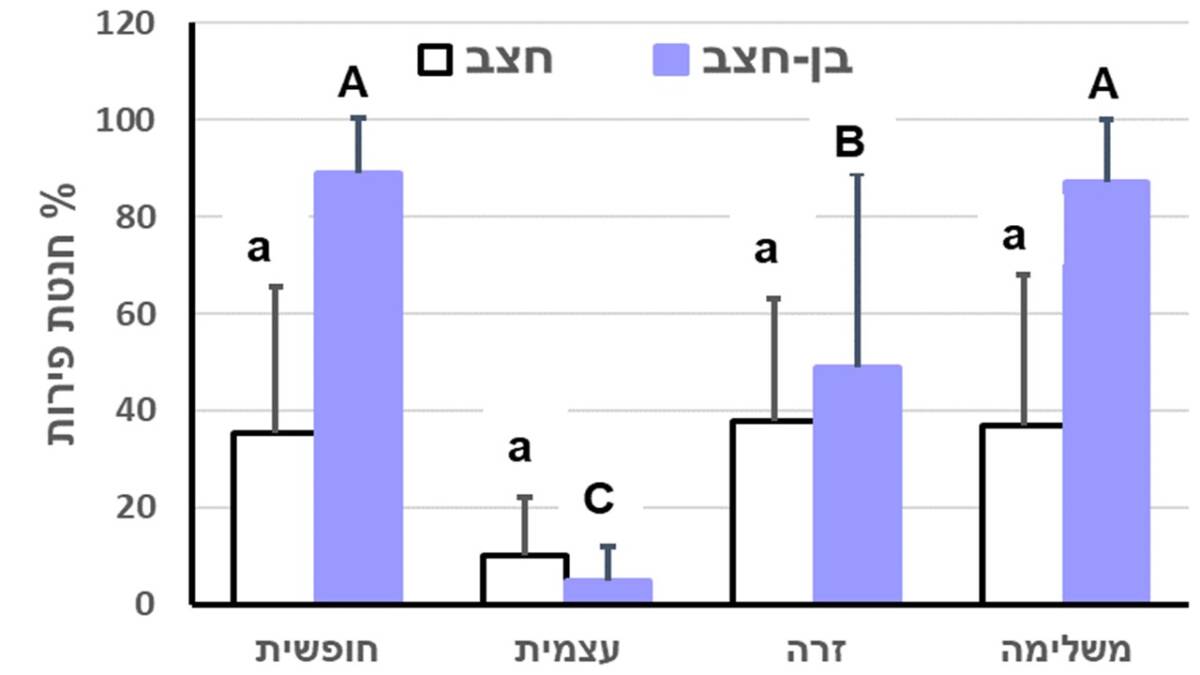
איור 3. אחוז חנטת הפירות (ממוצע ± סטיית תקן) בחצב מצוי ובבן-חצב יקינטוני לאחר האבקה חופשית, האבקה עצמית ספונטנית בפרחים מכוסים ברשת למניעת גישה של חרקים, האבקה זרה מלאכותית בפרחים מכוסים ברשת, והאבקה זרה משלימה בנוסף להאבקה חופשית. לגבי כל מין בנפרד, עמודות המסומנות באותיות זהות אינן נבדלות באופן מובהק (Duncan’s test P>0.05).
מספר הזרעים הממוצע (± סטיית תקן) בהלקט בפרחים שהואבקו באופן חופשי בחצב מצוי היה 9.33±3.58, ומספר הזרעים הממוצע (± סטיית תקן) בהלקט זה בבן-חצב יקינטוני היה 3.16±1.13 וההבדל היה מובהק (T(37.9)=9.699, P=0.0001). מספר הזרעים הממוצע (± סטיית תקן) שייצר צמח שלם של חצב מצוי היה 170.4±170.4 וזה שייצר צמח של בן-חצב יקינטוני היה 35.2±21.4, והבדל זה היה מובהק (T(33.8)=4.599, P=0.0001). מספר הזרעים להלקט לא הושפע מטיפולי ההאבקה השונים (איור 4).

איור 4. מספר זרעים בפרי (ממוצע ± סטיית תקן) בחצב מצוי ובבן-חצב יקינטוני לאחר האבקה חופשית, האבקה עצמית ספונטנית בפרחים מכוסים ברשת למניעת גישה של חרקים, האבקה זרה מלאכותית בפרחים מכוסים ברשת, והאבקה זרה משלימה בנוסף להאבקה חופשית. עמודות המסומנות באותיות זהות אינן נבדלות באופן מובהק (Duncan’s test P>0.05).
אבקה, ביציות, זרעים והצלחה רבייתית
מספר גרגרי האבקה המיוצרים על-ידי פרח (טבלה 1) היה נמוך במעט בבן חצב יקינטוני מאשר בחצב מצוי וההבדל היה גבולי אך מובהק (T(58.0)=7.28, P=0.4997). לעומת זאת מספר הביציות בפרח של חצב מצוי היה גבוה יותר והיחס O/P נמוך יותר בחצב מצוי יחסית לבן-חצב יקינטוני (טבלה 1) והבדלים אלה היו מובהקים (T(22.8)=54.5, P=0.0001) למספר ביציות, (T(42.9)=27.52, P=0.0001) ליחס O/P. אחוז הביציות שהופרו והפכו לזרעים (S/O) היה יותר מכפול בבן-חצב יקינטוני מאשר בחצב מצוי וההצלחה הרבייתית היחסית הייתה פי שש גבוהה יותר (RRS). מכיוון שהערכים חושבו מהמוצעים לכל מין, לא ניתן היה לבצע מבחן לבדיקת מובהקות ההבדלים.
השקעת ביומסה
בטבלה 2 מוצגים הערכים של הביומסה המושקעת באברי הצמח השונים של שני המינים הנחקרים. בשל ההבדל הגדול בביומסה הכוללת של שני המינים, ההשקעה בוטאה גם באחוזים מהביומסה הכללית. למרות הבצל הגדול יותר של חצב מצוי, ההשקעה היחסית בבצל הייתה נמוכה יותר מאשר בבן-חצב יקינטוני (טבלה 2). לעומת זאת ההשקעה היחסית בעלים, עמוד תפרחת ופרחים הייתה גבוהה יותר בחצב המצוי מאשר בבן-חצב יקינטוני. ההשקעה הממוצעת בעמוד התפרחת ובפרחים הדרושה לייצור זרע אחד הייתה 19.4 גרם בחצב מצוי ורק 0.84 גרם בבן-חצב יקינטוני.
דיון
הכשירות של צמח בסביבתו הטבעית מושפעת מתנאי הסביבה וממגוון תכונות ווגטטיביות (הקשורות לצמיחה) ורפרודוקטיביות (הקשורות לרבייה) שלו. לכן, בבואנו להשוות את הכשירות של שני מינים, רצוי מאוד לבדוק את המרכיבים השונים המשפיעים עליה. הצלחה רבייתית דומה של שני מינים יכולה להיות תוצאה של השקעה מוחלטת או יחסית, שונה בגורמים השונים המשפיעים עליה, כמו למשל זרעים קטנים ורבים כנגד מעט זרעים גדולים. לכן, במחקר זה השוויתי היבטים שונים של הביולוגיה של הרבייה וההשקעה של ביומסה בחלקי צמח בעלי תפקידים שונים. ההשוואה נערכה בין שני מינים דומים וקרובים, שבעבר אף נמנו על אותו סוג. השוואה מפורטת זו באה על מנת לבדוק האם ההסטה האבולוציונית של מועד הפריחה של חצב מצוי מהאביב לסתיו, קשורה ליתרון רבייתי כלשהו למין זה.
האבקה, חנטת פירות ויצור זרעים
אחוז חנטת הפירות באוכלוסיות טבעיות הוא גורם מרכזי המשפיע על ההצלחה הרבייתית של צמח, והוא מושפע מהשקעה מוגבלת של משאבים ובמקביל גם מפעילותם של מאביקים. תוצאות המחקר מראות כי אחוז חנטת הפירות בחצב מצוי בסתיו היה נמוך מאשר של בן-חצב יקינטוני באביב, דבר העשוי לשקף חסרון רבייתי. השונות הגבוהה מאוד באחוז החנטה, מרמזת על חנטת פירות נמוכה מהמיטבית, אולי בשל שונות במשאבים בבית גידול הטרוגני, או הבדלים ניכרים בפעילות המאביקים. הרמה הנמוכה מאוד של חנטת פירות בהאבקה עצמית ספונטנית (או בהאבקת רוח) מוכיחים שהרבייה בשני המינים תלויה באופן מוחלט, כמעט, בפעילותם של המאביקים.
חנטת פירות כתוצאה של האבקה מלאכותית זרה משלימה ,של פרחים שהיו חשופים לפעילות הרגילה של המאביקים בסביבה, יחסית לחנטת הפירות הטבעית יכולה לגלות האם פעילותם של המאביקים היא גורם המגביל לחנטת פירות או לחילופין משאבים מוגבלים הם הגורם המגביל (Stephanson1981, Zimmerman & Pyke 1988). האבקה משלימה הגורמת להגדלת אחוז החנטה של פירות היא עדות שפעילות המאביקים מגבילה את חנטת הפירות. לעומת זאת העדר השפעה של האבקה משלימה מוכיחה שמשאבים הם המגבילים את אחוזי החנטה. העובדה שהאבקה משלימה לא הגדילה את אחוז חנטת הפירות בחצב המצוי ובבן-חצב היקינטוני, מראה שבאביב וגם בסתיו יש פעילות מאביקים מספקת הגורמת למיצוי פוטנציאל הרבייה של כל אחד מהמינים הנבדקים (אבל ראו Young & Young 1992).
מספר הזרעים בהלקט היה גבוה יותר בחצב מצוי מאשר בבן-חצב יקינטוני, אבל הזרעים של חצב קטנים וקלים יותר מאלו של בן-חצב. הבדל זה עשוי לשקף הבדלים מוכרים באסטרטגיית הרבייה של מינים. מיני חלוץ למשל, ידועים כמייצרים זרעים רבים וקטנים בעוד צמחי קיימא מייצרים זרעים מעטים אך גדולים יותר (Moles & Westoby 2006). יתכן שהבדל זה משקף את ההבדל באסטרטגיות החיים והנביטה ואת ההבדל בבתי הגידול של שני המינים. חצב מצוי גדל בעיקר במקומות מופרים ושטופי שמש, בעוד בן-חצב יקינטוני גדל בעיקר בבתי גידול מוצלים יותר, בהם לזרע גדול יש יתרון יחסי להתבססות של נבטים. מספר הזרעים הנמוך בהלקט, בטיפולי ההאבקה העצמית המלאכותית, עשוי לשקף מצב של אי-סבילות עצמית, בו האבקה עצמית אינה מאפשרת הפרייה (איזיקוביץ 1970). חנטת פירות ועשיית זרעים הם מדדים של הצלחה רבייתית נקבית. העובדה שלא נמצאו הבדלים מובהקים בין טיפולי ההאבקה בחנטת הפירות ומספר הזרעים להלקט, במרבית המקרים, אינה פוסלת הבדלים בהצלחה רבייתית זכרית כפי שנמצא במינים אחרים (Queller 1984, Brunet 1992).
אבקה, ביציות, זרעים והצלחה רבייתית
היחס בין מספר גרגירי האבקה למספר הביציות (O/P) שמייצר פרט, או פרח במקרה של צמחים בעלי פרחים דו-מיניים, מהווה אינדיקציה טובה בקנה מידה אבולוציוני, למנגנון הזיווג ולהשקעה הזכרית או הנקבית בפרחים (Queller 1984).יחס זה, נותן מושג כמה גרגירי אבקה דרושים על מנת להפרות ביצית אחת. יחס O/P יורד לאורך גרדיינט בין מינים המחייבים האבקה זרה, לאלו הפוריים גם בהאבקה עצמית, ופרחים המתרבים באמצעות פרחים קלייסטוגמיים, שאינם נפתחים כלל, ומתבצעת בהם רק האבקה עצמית (Cruden 1977). יחס O/P משתנה עם שינויים בסינדרום ההאבקה, הוא גבוה בצמחים מואבקי רוח (בהם יש בזבוז גדול של אבקה) ונמוך בסינדרומים יעילים יותר של האבקה ביוטית על-ידי בעלי חיים המעבירים ביעילות אבקה מפרח לפרח, אבל הוא מושפע גם מגורמים נוספים (Dafni 1992). כשמשווים יחס זה בין צמחים של מינים קרובים, עם מבנה פרח וחברת מאביקים דומים, הוא יכול לשמש כאינדיקטור אמין ליעילות ההאבקה ולהצלחה זכרית או נקבית של המינים. לכן יחס O/P נמוך יותר בחצב מצוי עשוי לרמז על יעילות גבוהה של האבקה והצלחה זכרית גבוהים יותר יחסית לבן-חצב יקינטוני. תוצאות אלה אינן מתאימות להאבקת רוח שדווחה בעבר לגבי אוכלוסיות אחרות של חצב מצוי (Dafni & Dukas 1986).
אחוז חנטת הפירות ומספר הזרעים בהלקט מהווים שניהם מדדים להצלחה נקבית (Charlesworth 1989), והמכפלה שלהם משקפת את ההצלחה הנקבית היחסית הכללית של צמחים RRS (Wiens 1984). הערך, של כל המדדים האלה היה גבוה יותר בבן-חצב יקינטוני ומעיד על הצלחה נקבית גבוהה יותר מאשר של חצב מצוי. מכיוון שהצלחה נקבית מושפעת לעיתים קרובות מזמינות המשאבים, יתכן שהבדל זה משקף את זמינות המשאבים, במיוחד בתקופת הבשלת הפירות. זמינותם טובה יותר באביב בתקופת הפריחה והבשלת הפירות של בן-חצב יקינטוני מאשר בסתיו בזמן הפריחה והבשלת הפירות של החצב המצוי.
ביומסה
לחצב המצוי בצל גדול יותר מאשר זה של בן-חצב יקינטוני. עובדה זו מתיישבת עם שתי הנחות: 1. גאופיטים הגדלים בבתי גידול יובשניים ובלתי חזויים אוגרים יותר חומרי תשמורת מאשר לאלה הגדלים בבתי גידול נוחים וחזויים. 2. גאופיטים היסטראנתיים, הפורחים ללא עלים, אוגרים יותר חומרי תשמורת מאשר סינאנתיים הפורחים במקביל לעלים (Dafni 1981a). אבל אם מסתכלים על הנתונים המשוקללים, יחסית לביומסה הכללית של הצמח, התמונה משתנה. בן-חצב יקינטוני אוגר אחוז גבוה יותר מהביומסה הכללית של הצמח מאשר חצב מצוי. גם ההשקעה היחסית של ביומסה בזרעים גבוהה יותר בבן-חצב יקינטוני מאשר בחצב מצוי. מעניין שההשקעה היחסית בעלים הייתה נמוכה יותר בבן-חצב יקינטוני, דבר המרמז על פעילות הטמעה יעילה יותר (ליחידת ביומסה של עלה) מאשר בחצב מצוי.
ההשקעה בפרחים, למשיכת מאביקים בפרחים, נמוכה יותר ככל ששיעור ההאבקה העצמית הספונטנית גדולה יותר. כמו כן, ההשקעה באברים שונים בפרח היא יחסית לתרומתו של אותו חלק במשיכת מאביקים (Bell 1985). לכן, בצמחים הפורחים בזמן שיש תחרות חזקה על מאביקים, כפי שמצויה בישראל באביב, צפויה ההשקעה באיברים מושכי-מאביקים להיות גדולה יותר מאשר בצמחים הפורחים בזמן של תחרות נמוכה על מאביקים. בניגוד לכך, תוצאות המחקר מראות שההשקעה היחסית בעמוד התפרחת ובפרחים, הקשורים שניהם במשיכת מאביקים היו נמוכים יותר בבן-חצב יקינטוני (5.6%) הפורח באביב מאשר בחצב המצוי (16.8%) הפורח בסתיו. זאת, למרות שההשקעה היחסית בזרעים הייתה גדולה יותר בבן-חצב יקינטוני (4.6%) מאשר בחצב מצוי (2.6%). ההשקעה במשיכת מאביקים יעילה יותר בבן-חצב יקינטוני, למרות שהוא פורח באביב בזמן תחרות גבוהה על מאביקים, מאשר בחצב מצוי הפורח בסתיו בזמן של מיעוט פרחים מתחרים. אבל, למיטב ידיעתי,לא נערכה השוואה יסודית של עוצמת התחרות על מאביקים בין האביב לסתיו. השוואה שאמורה להתחשב במספר המאביקים פר-פרח בכל תקופה, ולקחת בחשבון גם את מגוון הפרחים ומגוון המאביקים, ולא רק את השפע הכללי שלהם.
סיכום
בשלב זה נוכל לסכם:
- האבקה אינה גורם מגביל לחנטת פירות בחצב מצוי הפורח בסתיו, עדות לכך שאין מחסור במאביקים בעונה זו.
- האבקה אינה גורם מגביל גם לחנטת פירות בבן-חצב יקינטוני הפורח באביב, עדות לכך שאין מחסור במאביקים גם בעונה זו.
- שיעור ההצלחה הזכרית ברביה, כפי שבאה לידי ביטוי ביחס O/P, גבוה יותר בחצב מצוי מאשר בבן-חצב יקינטוני.
- שיעור ההצלחה הנקבית ברבייה, כפי שמתבטאת ב RRS גבוה יותר בבן-חצב יקינטוני.
- ההשקעה היחסית של ביומסה למשיכת מאביקים, נמוכה יותר בבן חצב יקינטוני מאשר בחצב מצוי, אבל היא מספיקה להשגת שיעור הצלחה נקבית גבוה יותר.
התוצאות מראות יתרון בשיעור ההצלחה הזכרית של חצב מצוי הפורח בסתיו, זאת בהתאמה להשערת המחקר. אבל ההסטה של הפריחה מהאביב לסתיו הייתה, כנראה, כרוכה בהגדלת ההשקעה במשיכת המאביקים, וירידה בשיעור ההצלחה הנקבית. כל זאת, בתנאי שההשוואה של שני המינים שנחקרו, מייצגת נכונה את כלל הגאופיטים פורחי-האביב לעומת אלה הפורחים בסתיו.
גורם נוסף, שלא נבדק במחקר זה, הוא גורל הזרעים והנבטים של שני המינים. יתכן ששיעור טריפת הזרעים של חצב מצוי, החשופים לטריפה כחודשיים שלושה בלבד, נמוך יותר מאשר זה של הזרעים של בן-חצב יקינטוני הפורח באביב, וזרעיו מחכים בקרקע כחמישה חודשים, כשהם חשופים לטריפה. הבדל זה, אם יאושש במחקר, עשוי להוות יתרון גדול, שאולי הניע את הסטת מועד הפריחה של החצב מהאביב לסתיו.
ספרות
איזיקוביץ ד 1970 אקולוגיה של ההאבקה בצמחי חוף הים בישראל. תיזה לדוקטורט, אוניברסיטת תל אביב.
גליל י 1961 קינטיקה של גאופיטים. הקיבוץ המאוחד, תל-אביב.
Archibold OW 1995 Ecology of World Vegetation. Chapman & Hall, London.
Arroyo A & Dafni A 1995 Variation in habitat, season, flower traits and pollination in dimorphic Narcisus tazetta L. (Amarylliaceae) in Israel. Mew Phytologist 129:135-145.
Auerbuch M & shmida A 1987 Spatial scale and determinants of species richness. Trends in Ecology and Evolution 2:238-242.
Bell G 1985 On the function of flowers. Royal Society London B 224:223-265.
Bloom AJ Chapain FS & Mooney HA 1985 Resource limitation in plants – an economic analogy. Annual Review of Ecology and Evolution 16:363-392.
Brunet J 1992 Allocation in hermaphroditic plants. Trends in Ecology and Evolution 7: 79-84.
Campbell DR & Motten AF 1985 The mechanism of competition for pollination between to forest herbs. Ecology 66:554-563.
Charlesworth D 1989 Evolution of low female fertility in plants: pollen limitation, resource allocation and genetic load. Trends in Ecology and Evolution 4:289-292.
Cohen D & Shmida A 1993 The evolution of floral display. Evolutionary Biology 27:197-243.
Cruden RW 1977 Pollen ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31:32-46.
Dafni A 1992 Pollination Ecology a Practical Approach. Oxford University Press, Oxford.
Dafni A Cohen D & Noy-Meir I 1981a Life-cycle variation in geophytes. Annals on Missouri Botanical Gardens 68:652-660.
Dafni A & Dukas R 1986 Insect and wind pollination in Urginea maritima (Liliaceae). Plant Systematics and Evolution 154:1-10.
Dafni A Shmida A & Avishay M 1981b Leafless autumnal-flowering geophytes in the Mediterranean region – phytogeographical, ecological and evolutionary aspects. Plant Systematics and Evolution 137:181-193.
Enquist BJ West GB Charnov EL & Brown JH 1999 Allometric scaling of production and life-history variation in vascular plants. Nature 401:907-911.
Evenson WF 1983 Experimental studies of reproductive energy allocation in plants. In: Jones CE & Little RJ (editors) Handbook of Experimental Pollination Biology. Scientific and Academic Publications, New York pp:187-214.
Gentry AH 1974 Flowering phenology and diversity in tropical Bignoniaceae. Biotropica 6:64-68.
Heinrich B 1975 Energetics of pollination. Annual Review of Evolution and Systematics 6:139-170.
Heinrich B & Raven PH 1972 Energetics and pollination ecology Science 76:597-602.
Kochmer JP & Handel SN 1986 Constrains and competition in the evolution of flowering phenology. Ecological Monographs 56:303-325.
Kozlowski J 1992 Optimal allocation of resources to growth and reproduction: Implications for age and size at maturity. Trends in Ecology and Evolution 7:15-19.
Letty C Dyre RA Verdoor IC & Codd LE 1962 Wild flowers of the Transvaal. Centaur Publishers, Pretoria.
Moles AT Westoby M 2006 Seed size and plant strategy across the whole life cycle. Oikos 113:91-105.
Mosquin T 1971 Competition for pollinators as a stimulus for the evolution of flowering time. Oikos 22:398-402.
Orshan G 1983 Approches to the definition of Mediterranean growth forms. In: Kruger FJ Mitchel DT & Jarvus UM (editors) Mediterranean Type Ecosystems: The Role of Nutrients. Springer Verlag, Berlin.
Plants of the world online, Kewscience, Royal Botanic Gardens, Kew, http://powo.science.kew.org accessed 28.08.2020.
Pleasant JM 1980 Competition for bumblebee pollinatorsin Rocky mountain plant communities. Ecology 61:1446-1459.
Primack & Stacy E 1998 Cost of reproduction in the pink lady's slipper orchid (Cypripedium acaule, Orchidaceae): an eleven‐year experimental study of three populations. American Journal of Botany 85:1672-1679.
Queller DC 1984 Pollen-ovule ratios and hermaphrodite sexual allocation strategies. Evolution 38:1148-1151.
Raunkiaer C 1934 The Life Forms of Plants and Statistical Plant Geography. Clarendon Press, Oxford.
SAS Institute 1988 SAS/STAT User Guide. SAS Inc., Cary.
Shmida A & Dafni A 1989 Blooming strategies, flower size and advertising in the ‘lily group’ geophytes in Israel. Herbertia 45:11-123.
Schwartz-Tzachor R Eisikowitch & D Dafni A 2008 Flower characteristics and breeding system of two phenological ecotypes of Cyclamen persicum Mill. (Myrsinaceae) in Israel. Plant Systematics and Evolution 274:127-134.
Shon JJ & Policansky D 1977 The cost of reproduction in the mayapple Podophyllum peltatum (Berberidaceae). Ecology 58:1366-1374.
Stephanson AG 1981 Flower and fruit abortion: Proximate causes and ultimate functions. Annual Review of Ecology and Systematics 12:253-279.
Waser NM 1979 Pollinator availability as determinant of flowering time in ocotillo (Fouquieria splendens). Oecologia 39: 107-121.
Waser NM 1983 Competition for pollination and floral characteristics among sympatric plant species: A review of evidence. In: Jones CE & Little RJ (editors) Handbook of Experimental Pollination Biology. Scientific and Academic Editions, New York pp:187-214.
Wiens D 1984 Ovule survivorship, brood size, life history, breeding systems and reproductive success in plants. Oecologia 64:47-53.
Willson MF 1983 Plant Reproductive Ecology. John Wiley & Sons, New York.
Young HJ & Young TP 1992 Alternative outcome of natural and experimental high pollen loads. Ecology 73:639-647.
Zimmerman M & Pyke GH 1988 Reproduction in Poleminium: Assessing the factors limiting seed set. American Naturalist 131:723-738.
Zohary M 1976 Plant Life in Palestine. Ronald Press, New York.
==============================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: נאמן ג 2020 למה החצב המצוי פורח בסתיו? השוואת ההשקעה בביומסה וברביה, בינו לקרובו בן-חצב יקינטוני הפורח באביב. כתב-עת "כלנית", מספר 7.










לפוסט הזה יש 2 תגובות
שלום
למיטב ידיעתי החצב הגיע אלינו מדרום אפריקה, שם הוא פורח באותו תאריך כמו בישראל כלומר שם באביב.
זה אחד מהמקרים שצמח שהגיע מחצע הכדור הדרומי. לא שמר על עונת הפריחה כאשר עבר לחצי הצפוני. אלא שמר על התאריך והפח מצמח פורח אביב לצמח פורח סתיו.
ואס כל המחקר היסודי והמענין משתבש.
אשמח לתגובה 0547916602.
תודה ושנה טובה
שלום,
מניין הידע על מוצא החצב בדרום אפריקה ונדידתו או יבוא שלו לישראל? בכל מקרה הוא נציג בכיר של קבוצת גאופיטים הפורחים בסתיו טרם הגשמים.
גידי נאמן