

אורה הורוויץ ז"ל – המכון למדעי החיים, האוניברסיטה העברית ירושלים
פורסם מתוך עזבונה של אורה הורוויץ
עיבדו, אספו חומרים והביאו לדפוס – עוזי כוכבא ואבי שמידע
תקציר: אלה חומרים אשר היו בעזבונה של אורה הורוויץ זכרונה לברכה (2017-1920), אשר לא זכתה להשלים כתב-יד כוללני עברי אודות הביולוגיה והתירבות של הכלנית המצויה Anemone coronaria. מאז שנות השישים של המאה הקודמת עבדה אורה על האקולוגיה והגנטיקה של הכלנית המצויה והתרכזה בלימוד הזנים הצבעונים של המין, תפוצתם הגיאוגרפית, הגנטיקה של צבעי העטיף ואסטרטגיות האבקת-רוח והאבקת חרקים בכלניות האדומות לעומת הזנים הלא האדומים. ביחד עם חקלאי מושב זכריה טפחה כבר לפני חמישים שנה זנים תרבותיים צבעוניים של הכלנית ותרמה רבות להפיכתה של הכלנית "לפרח הלאומי של ישראל".
===============
סקירה טקסונומית
הסוג כלנית Anemone שייך למשפחת הנוריתיים ומקיף כ- 70 מינים עשבוניים הנמצאים בעיקר בחצי הכדור הצפוני, בעולם הישן ובעולם החדש – כאחד. בתוך הסוג, הכלנית המצויה .A. coronaria L, שייכת לסקצית כלנית Sec. Anemone [תיקון שנתתי במאמר מ- 1975 לסיווג הסיסטמטי שנתן אולבריך (Ulbrich) בתוך הסוג כלנית (Horovitz,1975)], מספר הכרומוזומים בתאי הכלנית המצויה הוא 2n=16, כלומר 8 זוגות.
המין גדל מהרי טיין-שאן במרכז אסיה דרך טורקיסטן, דרום איראן, עיראק, סוריה, לבנון, ירדן, ישראל, מצרים, לאורך חופי צפון-אפריקה ומערבה עד לתוניס ולאלג’יריה. הוא אינו גדל במרוקו, אך התפשט במסלול צפוני דרך ארמניה, טורקיה, הבלקן, חצי האי האיטלקי, דרום-מזרח ודרום צרפת, ספרד מערבה עד לגיברלטר. מין זה מהווה גשר סיסטמטי-ביוגיאוגרפי בין טווח רחב של מיני כלנית אסיאתיים (ראה- Floras of Russia) ביניהם מין הכלנית הקרוב ביותר כלנית ביפלורה Anemone biflora (מין ויקרי לכ. מצויה שלו תפוצה רחבה מטורקיסטן ועד פרס), וקבוצה הכלניות הים-תיכונית Anemone palmata, אשר חבריה העיקריים הינם A. pavonina ו- A. horentsis וצורות המשנה של מין זה. אף לא אחד מקרובים אלה גדל בישראל. השוני העיקרי בין כלנית מצויה למין כלנית ביפלורה הוא: לכלנית ביפלורה יש תפרחת בת 2-3 פרחים והעטיף שלו מתקשה כדי להגן על ראש הזרעים. מאידך שונה הכלנית המצויה מכלניות קבוצת פלמטה .A. palmata gr הגדלות מערבה לה, בכך שתפרחתה הצטמצמה לפרח בודד, והעטיף אשר מתפקד כאבר פרסומת הינו כותרתי לחלוטין, כלומר חסר עלי גביע. לכלנית המצויה שונות רבה בגודל הצמח, בטיפוסי העלים, בצבעי הפרחים ובצורתם, והיא פוצלה על ידי החוקרים אשר עסקו בטיפוח טיפוסי כלניות תרבותיות למינים שניתנים להכלאה מלאה לעומת כאלה שלא. על כן בזנים התרבותיים של הכלנית המצויה לא ניתן עוד להכיר את מין המקור.
 פולימורפיזם בכלנית מצויה Anemone coronaria , ראש העין, צילם עוז גולן ©
פולימורפיזם בכלנית מצויה Anemone coronaria , ראש העין, צילם עוז גולן ©
קשרים סיסטמטיים והכלאה בין בין מינים קרובים בקומפלקס כלנית מצויה
ייתכן שקיימת שאלה בדבר המעמד הסיסטמטי של חלק מן הטקסונים של מיני הכלנית במרכז אסיה כגון: Anemone eranthoides, A. regel, A. almaatenis A. seravshanica ושווה לבחון זאת באופן ניסויי. אינני יודעת דבר בעניין זה. הכלנית המצויה, בתפוצתה הים-תיכונית, סימפטרית (כלומר בעלת חפיפה גיאוגרפית) לעיתים עם מינים מקבוצת A. palmata, אך לא ראיתי דיווחים על מכלואים. חריג אחד הינן תצפיות שפורסמו בשנת 1883 על ידי Abbé A. Pons, בהן הוא תיאר מכלואים משוערים בין A. hortensis לבין כלנית מצויה ליד ניס שבריוויירה הצרפתית. כפי הנראה, לחלק מן הצמחים היו אבקנים מנוונים בעלי מאבקים מחודדים, דהיינו, היו עקרים זכרית. זה מתאים לתופעה של חוסר התאמה בין ציטופלסמה של הורה אחד לגנים גרעיניים מחזירים Restorer gene של ההורה האחר. כלומר הגן גרעיני מחזיר את הצמח לתפקודו הרגיל אשר נפגע כתוצאה מפגם בגן המיטוכונדרי; בדרך כלל מדובר בגן המחזיר פוריות לצמח עקר זכר. בכלנית ידוע היטב מכלוא בין מיני הכלנית A. hortensis לבין A. pavonina, המוכר בשם כ- A. x fulgenes, המבסס את עצמו באזורים בהם שני ההורים גדלים בחפיפה גיאוגרפית. דור כלאים F1 זה מגודל גם כצמח נוי ומופיע בקטלוגים של גינון.



מימין: כלנית מצויה ורודה ("פסטל"בלשון אורה) גדלה בתוך מרבד של כלניות אדומות ביער שוהם; צבעה נגרם על ידי צרוף הגנים Sc–,Ww,B (minus) [ראה מקרא באיור בהמשך]. במרכז: כלנית מצויה אדומה Anemone coronaria. שתי התמונות צולמו על ידי גילה אלון. משמאל: רוב מיני הכלניות בעולם גדלות ביערות ממוזגים ברחבי אירו-אסיה וצבען לבן או כחלחל כמו מין כלנית זו שצולם ביערות סין, הצלם מרינוס ורגר ©.



תפוצה בישראל
המין הנפוץ בישראל כלנית מצויה A. coronaria L. גדל באופן נרחב ורציף בשטחים פתוחים שטופי שמש מגבול לבנון ועד הנגב הצפוני, תוך שונות גדולה בצפיפות האוכלוסיות. מאז שראיתי אותן לראשונה בתחילת שנות ה-60 הפכו האוכלוסיות למקוטעות יותר ויותר, זאת בעקבות בנית בתים וסלילת דרכים.
 תפוצת הכלנית המצויה Anemone coronaria (כל הצבעים) בשנים 1980 – 2000 על פי תצפיות מרכז רתם, התצפיות של פרטים פורחים סוכמו כל חמש שנים. נעשה ע"י נ.לוין וא.שמידע 2007 , המעבדה לחישה מרחוק, המחלקה לגיאוגרפיה ©.
תפוצת הכלנית המצויה Anemone coronaria (כל הצבעים) בשנים 1980 – 2000 על פי תצפיות מרכז רתם, התצפיות של פרטים פורחים סוכמו כל חמש שנים. נעשה ע"י נ.לוין וא.שמידע 2007 , המעבדה לחישה מרחוק, המחלקה לגיאוגרפיה ©.

שונות גנטית באוכלוסיות כלנית מצויה בישראל
מקצת השונות של התכונות המורפולוגיות באוכלוסיות הכלנית המצויה היא קלינלית, כלומר משתנה בהדרגה לאורך גרדיאנט היובש הגיאוגרפי מצפון לדרום בישראל. שונות קלינלית זו משפיעה על אוכלוסיות שלמות. זה ניכר במיוחד בתכונות העלים, החלוקה המשנית של טרפי העלה ושעירותם גדלה לאורך גראדיאנט הצחיחות ממערב למזרח ומצפון לדרום. זמן הפריחה משתנה אף הוא לאורך גראדיאנט האקלים, הפריחה מקדימה בחלקים הקרים של המדינה, ופריחת סוף-עונה היא בחלקים החמים שלה (Horovitz 1975), אך יש גם מספר חריגות (ראה להלן).

פולימורפיזם בצורת-העלה ובפסיג של כלנית מצויה Anemone coronaria. האיור לקוח מתוך כרזה שאורה הורוויץ הציגה בכנס ב 1999 ©.
| הערה לגבי למועד הפריחה
הזן הלבן והזן הכחול של הכלנית המצויה מקדימים לפרוח בממוצע כ 12 ימים לפני הזן האדום. כיוון שהזנים הכחול והלבן גדלים בצפון ישראל, הכלילה אורה כי "הפריחה מוקדמת יותר באזורים הקרים" אך לא כן הדגם בתוך כל צבע. כך, הכלניות האדומות מקדימות לפרוח במעונות הנמוכים והחמים ומאחרות במעונות הגבוהים והקרים. למשל שיא הפריחה כלניות באתר שוקדה בנגב המערבי חל בממוצע בתאריך שבעה בפברואר, בעוד שיא הפריחה של הכלניות בירושלים הינו באחד במרץ, ובהר-מירון שיא הפריחה הינו במחצית חודש מרץ. |
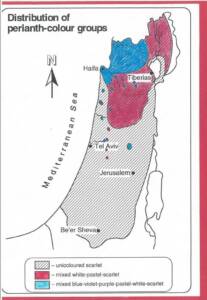

מימין: מפת תפוצת צבעי העטיף (הכותרת) בכלנית מצויה Anemone coronaria בישראל. מתוך כרזה שהציגה אורה הורוויץ בכנס ב 1999. משמאל: פקעת ענפה בעלת חמש אונות של כלנית מצויה, מתוך עזבונה של אורה הורוויץ ©.


צרוף של גורמי קרקע ואקלים קובעים את הרכב צבעי הפרחים באוכלוסיות השונות, כך שתחום המאוכלס על ידי כלנית מצויה ניתן לחלוקה לאזורים על פי צבעי הפרחים: כל הצבעים האפשריים; אדום ולבן עם מעט וורוד בהיר ואדום בלבד. קיים חומר רב בנושא זה במאמרי המוקדמים, המבוססים על עבודתי עם דניאל זוהרי, כסטודנטית שלו (Horovitz 1975, 1976).
שונות על בסיס מקומי שמשפיעה על כלל האוכלוסיה, אך ללא עדות לתבנית יסוד, ניתן לראות בגודל הצמח, בגודל הפרח ובגודל הפרי, במורפולוגיה של הפקעת ובמורפולוגיה של הפסיגים. אוכלוסיות בודדות מאופיינות לעתים על ידי צברי מוטנטים המציגים תכונת פרח נדירה כמו צבעי עטיף של מוטנטים נדירים. באיור בהמשך מוצגים מספר אתרים חשובים בישראל בהם נמצאו מקבצים נדירים של כלניות בעלי תכונות פנוטיפיות מיוחדות של צבע וכן של פרטים בעלי שניים או שלושה פרחים על גבעול תפרחת.
במחקרים שבוצעו ע"י יונש וחבריו (Yonash et al. 2004 וראה שם הפניה למאמרים נוספים) נמצא כי מידת השונות הגרעינית הנראית באמצעות AFLP Amplified Fragment Length Polymorphism) נבדלת באוכלוסיות ישראליות שונות.
 מוטנטים נדירים בצבעי העטיף (כותרת) בכלנית המצויה Anemone coronaria . מתוך כרזה אשר הציגה א.הורוויץ בשנת 1999 ©.
מוטנטים נדירים בצבעי העטיף (כותרת) בכלנית המצויה Anemone coronaria . מתוך כרזה אשר הציגה א.הורוויץ בשנת 1999 ©.
 מפת האתרים בהם נמצאו מקבצים נדירים של כלניות בעלי תכונות פנוטיפיות מיוחדות של צבע ומספר פרחים על גבעול . מתוך כרזה שהציגה א.הורוויץ ©.
מפת האתרים בהם נמצאו מקבצים נדירים של כלניות בעלי תכונות פנוטיפיות מיוחדות של צבע ומספר פרחים על גבעול . מתוך כרזה שהציגה א.הורוויץ ©.

שונות גנטית מיטוכונדרית
שונות גנטית מיטוכונדרית בכלנית המצויה נחקרה בארבע אוכלוסיות טבעיות של הכלנית המצויה בישראל באמצעות שיטת הקיטוע המבדיל RFLP (כוכבא 1998). אוכלוסיות הכלנית שנחקרו גדלו במרחק של 80-150 ק"מ זו מזו בבתי גידול שונים והיו שונות זו מזו בתכונות מורפולוגיות ופיזיולוגיות. בד בבד עם האוכלוסיות הישראליות נדגימו שתי אוכלוסיות בר של כלנית מצויה מאנטוליה בטורקיה, זני תרבות, הכלאות זן-בר x זן-תרבותי, נגזרות מכלוא ופרטים בודדים מהמין A. blanda .
החומר הגנטי המורכב מרצפי DNA מצוי בצמחים ובחיות בגרעין אך גם במיטוכונדריות ובכלורופלסטים. לעומת בעלי-חיים, בצמחים מולקולת ה DNA במיטוכונדריה יוצרת טבעת ענקית מפותלת הנראית בהגדלה כמו שער פרוע. בנוסף לטבעת העיקרית נמצאו עוד טבעות קטנות. הטבעת העיקרית התפתחה כנראה לשרשרת כה מוארכת ומפותלת בעקבות רקומבינציות רבות אשר גרמו לחיתוכים, היפוכים והכפלות של קטעי DNA. העבודה התמקדה בבדיקת השונות של הגנום המיטוכונדרי: שני אנזימי קיטוע (HindIII, EcoRI) שימשו לחיתוך DNA כללי, ותוצרי החיתוך הגיבו עם שלושה גלאים מיטוכונדריים (nad3, rps12, atpA). שימוש במערכת ההדמיה הממוחשבת phosphor analyzer הביא לזיהוי עוצמות תגובה של קטיעי DNA ברמות שונות, ולכימותם. מתוך 83 צמחים שנבדקו בששה צירופי אנזים/גלאי התקבלו 20 קטעים שיצרו שמונה מיטוטיפים שונים כאשר הקטיעים בעלי עוצמת התגובה המירבית בלבד נלקחו בחשבון, לעומת 62 קטיעים ו- 47 מיטוטיפים שהתקבלו כאשר כל הקטיעים נלקחו בחשבון. כל קבוצות הכלנית שנבדקו היו רב צורתיות. רק הצרוף EcoRI/atpA נמצא חד-צורתי ב- A. coronaria.

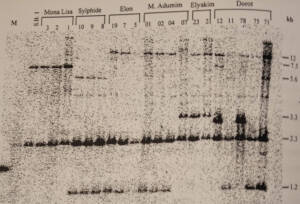
מימין: מידת השונות התוך אוכלוסייתית בישראל כפי שהיא מתבטאת בהרצת קטעי DNA בג'ל; הכתמים הכהים מציינים אזורים המגיבים לגנים מיטוכונדריים מסומנים ראדיואקטיביים, מעובד מתוך ע.כוכבא 1998. משמאל: מידת השונות באוכלוסיות בר לעומת זני תרבות של כלנית מצויה כפי שמתבטאת בעבודת ע.כוכבא 1998. הכתמים הכהים מציינים אזורים המגיבים לגנים מיטוכונדריים מסומנים ראדיואקטיביים. מעובד מתוך ע.כוכבא 1998 ©.
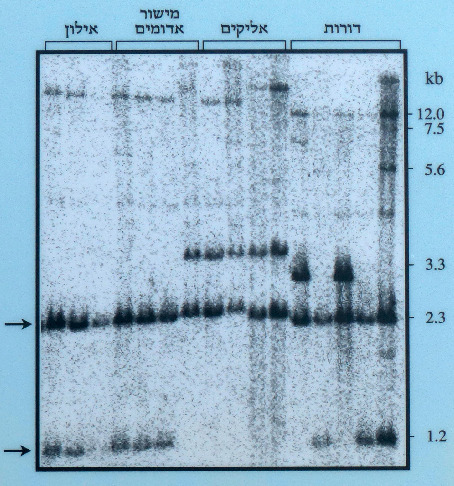

השונות באוכלוסיות הבר הישראליות, המבוססת על תדירות המיטוטיפים באוכלוסיה, Hs, היתה כ- 0.5 בהתבסס על הקטיעים בעלי עוצמת התגובה המירבית בלבד. מדד זה הגיע ל- 0.9 כאשר כל הקטיעים (קטע DNA הנחתך ע"י אנזימי חיתוך) נלקחו בחשבון, ובבדיקה זאת עברה השונות האוכלוסייתית הממוצעת Hs(m) את השונות בכלל אוכלוסיות ישראל Ht. עיקר השונות בישראל היא שונות תוך-אוכלוסייתית, והיא גדולה פי שלושה מהשונות הבין-אוכלוסייתית. הפרמטר θ של Weir, המתבסס על תדירות הקטעים באוכלוסייה, שימש למדידת ההתמיינות בין האוכלוסיות. ערך θ שנמדד עבור אוכלוסיה אחת מאליקים, היה פי עשרה גדול מזה שנמדד עבור שלוש האוכלוסיות האחרות, ודומה לערך המצביע על התמיינות האוכלוסיות הישראליות מאלה התורכיות (θ=0.31). נמצאו הבדלים ברורים בין דגמי הקטיעים של האוכלוסיות הישראליות לבין אלה של הצמחים מתורכיה, ונמצאה אבחנה ברורה בין כלל דגמי צמחי הבר לדגמי הקטיעים שהתקבלו מזני התרבות. מהשוואת המיטוטיפים של הורים וצאצאים עלה כי דגם הורשת הגנים המיטוכונדריים ב- A. coronaria הינו אימהי.
בניגוד למיני הצמחים שנחקרו לפני עבודה זאת, קיים בכלנית המצויה רק עותק אחד של הגן המיטוכונדרי atpA, בעוד שהגנים nad3 ו- rps12 קיימים במספר עותקים, חלקם ברמות תת-סטויכיומטריות שלא ניתנות למדידה, כפי שנמצא קודם לכן רק בטבק Nicotinia sylvestris. שני הגנים נמצאים במולקולה העיקרית של הכלנית המצויה בסמיכות רבה זה לזה, כפי שנמצא במינים אחרים, אך ייתכן כי במולקולות האחרות קיים רק אחד משני גנים אלה.
בקרה גנטית של תכונת צבע העטיף, מועד הפריחה וקשר לטיב הקרקע
| הסבר למושגים:
אלל (allele) – צורות שונות (כלומר הבדלים ברצף נוקלאוטידים) של גן. אחת מהגרסאות החלופיות של רצף DNA הנמצאת באתר גנטי מסוים. כל גן קיים בשני אללים (אלל אחד התקבל מכל הורה) אשר נמצאים בדיוק באותו מיקום על גבי כל אחד מהכרומוזומים. האללים יכולים להיות זהים או שונים זה מזה. לדוגמה לגן לצבע עיניים קיים אלל הקובע צבע כחול ואלל הקובע צבע חום. ביטוי התכונה תלוי ביחסי דומיננטיות-רצסיביות בין אללים של גן מסוים. רצסיבי (Recessive) – אחד משני אללים אשר אינו בא לידי ביטוי בפנוטיפ של היצור כאשר האלל השני הוא דומיננטי. מתייחס גם ליצור שנושא רק אלל/ים רצסיבי/ים לתכונה מסוימת. דומיננטי (Dominant) – כאשר אלל של גן ממסך באופן מלא את התבטאותו של אלל אחר ועותק יחיד ממנו מספיק כדי לגרום לפנוטיפ אליו הוא מקודד. הטרוזיגוט (Heterozygote) – יצור שיש לו שני אללים שונים של אותו גן בשני כרומוזומים הומולוגים. הומוזיגוט (Homozygote) – יצור שיש לו שני אללים זהים של אותו גן בשני כרומוזומים הומולוגים. מיטוטיפים (Mitotypes) – סדרת אללים של גנים מיטוכונדריים המורשים על ידי הורה אחד. |


מימין: סימול גנטי לצבעי הכלנית; בפוסטר אשר הציגה א.הורוויץ היא קשרה בין צבעי הכלנית השכיחים לבין המערכת הגנטית המפקחת על יצירת הצבע המסוים. בצבע "פסטל" התכוונה אורה לגוון עדין של ורוד או סגול המתווסף לכלניות לבנות. משמאל: סימולים גנטיים לצבעי הכלנית; מקרא לסימון הגנטי של גנים ואללים באיור 9א ובמאמר. אות גדולה-אלל דומיננטי; אות קטנה-אלל רצסיבי. צבעי פסטל: הכוונה לגוונים עדינים של ורוד וסגול על רקע העטיף הלבן. הסימול (-) מייצג אלל כלשהוא, לדוגמא הסימול (W-) משמעותו: בנוסף לאלל הדומיננטי W לא משנה איזה אלל יצטרף. מעובד מתוך ע.כוכבא 1998 ©.


חקרתי בכלנית המצויה אספקטים שונים אקולוגיה, ביולוגיה וגנטיקה, אולם משום שמוקד הקשר שלי עם הכלנית המצויה היה טיפוח פרחי קטיף, לכן מחקר צבע הפרחים היה נושא מרכזי (Horovitz 1975, 1995). צבעי העטיף (כותרת) של פרחי הכלניות מושג באמצעות תערובת של צבענים (פיגמנטים) פלאבונואידים, באמצעות ניסויי טיפוח של הכלאות פנוטיפים בעלי צבעים שונים הצלחנו למצוא את הגנים האחראים על בקרת הצבע. מתברר כי ארבעה גנים פועלים הדדית באופן אפיסטטי, הממסך אחד את משנהו (Horovitz 1975, 1995):
- הגן בצרוף האללים albalb (עבור albino = לבקן) חוסם התפתחות אנטוציאנין בכל חלקי הצמח ועל ידי כך מתקבל פרח אלבינו (לבן) לחלוטין. נדגיש כי בשונה מהגן לצבע לבן בעטיף הפרח, הגן ללבקנות alb הוא אפיסטטי על כל שאר הגנים, כלומר גורם להלבנה של כל אברי הפרח כולל האבקנים.
- הגן בצרוף האללים scsc (עבור scarlet = אדום) חוסם התפתחות צבעים שונים מאדום והופעתו גורמת לצבע עטיף אדום בלבד. בגנטיקה מקובל לסימון "Sc" אלל דומננטי ו "sc" אלל רצסיבי. המקום של הגן לאורך הכרומוזום קרוי "לוקוס" ומסומן בסימול -" SC/sc".
- הגן בצרוף האללים ww (עבור w = לבן) חוסם התפתחות צבעים לא-אדומים פרט ללבן.
- הגן בצרוף האללים bb בנוכחות האלל W מקודד עבור סגול וכחול.
הגנוטיפ scsc המתבטא בפנוטיפ עם פרח אדום הינו השכיח ביותר בישראל, בעוד שהגנוטיפ Sc-W-bb הוא כנראה השכיח בלבנון. נראה לי שזהו למעשה הגנוטיפ WWbb, כי מצאתי שהאלל w חסר באוכלוסיות הסמוכות לגבול לבנון. באוכלוסיות אדומות צפופות מאוד מצאתי שני אללים נוספים בלוקוס SC/sc, שניהם רצסיביים לאלל sc. הטיפוס ההומוזיגוט sc1sc1 מקודד לצבע פרח וורוד-תות והטיפוס ההומוזיגוטי sc2sc2 מקודד לצללית כמעט-כתומה (Horovitz 1979).
תחום הגנטיקה של צבעי הכלנית הוא תחום קל המתאים לתרגילי מעבדה בביולוגיה, כמו למשל: סטודנטים יכולים לזהות גנוטיפ הטרוזיגוטי Scsc באמצעות נקודות אדומות המופיעות על עטיף לא-אדום, לרוב עטיף לבן (Horovitz 1966). ניתן היה לשער שזיהוי גנוטיפ זה הוא חסר ערך, מכיוון שצבעני הפרח הינם תוצרים מטבוליים מבוזבזים. אך, מתברר כי לוקוס הגן Sc/sc מבקר ומשפיע לפחות על שתי תכונות נוספות בצמח, דרישות קרקע ומועד הפריחה. התברר לנו כי לצמחים הנושאים את האלל Sc יש דרישות קרקע מיוחדות בלעדיות, ושצמחים החסרים אלל זה והם בעלי גנוטיפ הומוזיגוטי scsc פורחים מאוחר יותר (Horovitz 1975). ניתן להבחין בזאת באוכלוסיות בר שבהן הפריחה מתחילה בפריחה מרובת צבעים, ההופכת מאוחר יותר בעונה לפריחה אדומה בלבד (Horovitz 1975). דגם זה אובחן גם ב"צאצאים מתפצלים" שנבדקו בניסיונות של הפריה מלאכותית בין גנוטיפים שונים. תוצאות אלה מצביעות כי לפחות אחד מן הגנים המוסיפים (Addetive genes) שמבקרים את תאריך הפריחה קשור לאלל sc. פריחה מוקדמת הינה דומיננטית על פריחה מאוחרת, תוצאה זו התקבלה גם בתצפיות על כלניות תרבותיות.
 פס וכתמי צבע אדום על עטיף כלניות לבנות. מעובד מתוך עזבונה של א.הורוויץ ©.
פס וכתמי צבע אדום על עטיף כלניות לבנות. מעובד מתוך עזבונה של א.הורוויץ ©.
כבר בשנים הראשונות לעבודתי בכלניות שמתי לב לכתמי צבע אדומים, אשר לעיתים מופיעים בצורת פס אדום על רקע העטיף הלבן של כלניות לבנות (Horovitz 1966). בהמשך. במחקר גנטי של הכלאות מלאכותיות התבררה התמונה: גנוטיפ הפרח הלבן הוא Scsc-ww כלומר הצבע הלבן ההומוזיגוטי בא לידי ביטוי כיוון שהגן לצבע האדום לא חוסם אותו. הצבע האדום מתבטא רק במצב ההומוזיגוטי רצסיבי scsc. כאשר הצמח הינו הטרוזיגוט, כלומר קיים בו האלל הדומיננטי Sc הצבע האדום איננו מופיע כלל בפנוטיפ. כך הוא המצב בגנוטיפ של רוב הכלניות הלבנות השכיחות ברחבי מרכז וצפון ישראל; העטיף הלבן הוא בעל גנוטיפ Scsc-ww אשר בו מתבטא הגן ww היוצר בפנוטיפ צבע לבן. אך במקרה שלפנינו, בזמן התפתחות ניצן הפרח תא אחד מרקמת העטיף איבד מסיבה כלשהיא את האלל הדומיננטי Sc, ולכן נותר רק אלל אחד שמשפיע על האדום האלל sc. במצב זה הגן לאדום הינו 'דמוי הומוזיגוט רצסיבי' והצבע האדום מתבטא והתא נצבע אדום. תוך כדי התפתחות ניצן הפרח התחלק התא האדום המוטנטי הזה לתאים רבים אשר יצרו צבר עגול של תאים אדומים או טורי תאים הוא הפס אדום הנראה בתמונות הנלוות.
 פס אדום בכלנית לבנה הנובע מ"פגם" בגן Scsc כאשר האלל Sc איננו מתפקד ובשל כך מתבטא האלל sc הרצסיבי והתא הופך אדום ו"צאצאיו" אשר התרבו בצורת פס יצרו פס אדום בעטיף הכלנית הלבן, צילמה גילה אלון ביער שוהם ©.
פס אדום בכלנית לבנה הנובע מ"פגם" בגן Scsc כאשר האלל Sc איננו מתפקד ובשל כך מתבטא האלל sc הרצסיבי והתא הופך אדום ו"צאצאיו" אשר התרבו בצורת פס יצרו פס אדום בעטיף הכלנית הלבן, צילמה גילה אלון ביער שוהם ©.
מגבלות טיפוח בתוך הכלנית המצויה בישראל
ביצעתי הרבה הכלאות צולבות בין אוכלוסיות ישראליות שונות (השונות בתנאי בית-הגידול, כמו גם בהרכב גנטי, בזמני פריחה ועוד) וכן בין אוכלוסיות ישראליות לאוכלוסיות מערב-תורכיות ובין אוכלוסיות ישראליות לזני-תרבות (בעיקר ממקור תורכי). כל צאצאי ההכלאות הללו התפתחו יפה בתנאי משתלה וגידול ישראלים. עם זאת, לא נבדקו מידת הצלחתם של תוצרי הכלאות אלה (רקומביננטים חדשים) בתנאי גידול טבעיים או בשתילה בתנאי בר.
להלן דוגמא אחת לאמירתי כי ההצלחה בתנאי משתלה איננה גוררת הצלחה בתנאי שדה: ידוע כי בעוד הפנוטיפים אדומי הפרח גדלים בכל סוגי הקרקעות, תערובות טיפוסים לא-אדומות של כחולים, וורודים ולבנים דורשות קרקעות עשירות במינרלים; בטבע כלניות כחולות ורדרדות ולבנות אלה אינן גדלות על קרקעות רנדזינה, חול או טרה-רוסה מסוננת (Horovitz 1975). באזורי מגע בין שני סוגי קרקעות, שמתרחש עקב פגם בסלע האם, גרגרי אבקה הנושאים את האלל הדומיננטי ’לא-אדום’ וגם זרעי צמחים לא-אדומים מופצים מקרקעות עשירות במינרלים אל קרקעות סמוכות עניות במינרלים, אולם אף לא צמח 'לא אדום' אחד אינו מצליח להגיע לשלב הריחה בקרקעות עניות במינרלים אלה.
מחזור חיים ופנלוגיה
באזור תפוצתם הים-תיכוני יש לצמחים משך חיים של 8-10 שנים ותקופת צעירות (יובניליות) של עד שלוש שנים. תרדמת-קיץ מתרחשת מידי שנה ונקבעת על פי טמפרטורה ואורך-יום. חלקי הצמח הנמצאים בתרדמת-קיץ הינם ניצנים אדוונטיביים חיצוניים על הפקעת. חלקים זקנים יותר של הפקעת הינם בני-חלוף. צאצאים של זרע שנשלח לדרום-אפריקה שמרו על פריחה בחורף, אך צאצאים של צמחים שעברו אקלום בצפון-מערב אירופה ובני תרבות שהתרבו מהם ושגודלו באזורים ממוזגים נמצאים בתרדמת-חורף ופורחים בקיץ. אין ספק כי תגובה לאורך יום ולטמפרטורה ניתנת למניפולציה בקלות (קדמן-זהבי והורוויץ 1980). קיימת שונות רבה בזמני הפריחה ובפיזור הזרעים, ועונת הפריחה הכללית היא נובמבר עד מרץ. על מנת לנבוט ולפתח פרי בעל זרע פורה נדרשים מספר חודשי המתנה לאחר ההבשלה, תכונה שהלכה לאיבוד בזני התרבות.
מערכת הזיווג mating system
עיקר ההאבקה בכלנית המצויה הינה האבקה זרה, ומנגנון האבקה זה נעזר בדיכוגמיה, היינו הפרדה בזמן ההבשלה של אברי הזכר מזה של אברי הנקבה. הכלנית ידועה כצמח פרוטוגיני, כלומר כזה שאברי הנקבה מבשילים בפרח טרם הבשלת אברי הזכר, זאת כדי למנוע האבקה-עצמית (Horovitz. 1975). מיד לאחר פתיחת הפרח הצלקות מוכנות לקליטת אבקה למשך מספר ימים מועט ואז ישנו פער של 8-12 ימים לפני שהמאבק הראשון מפזר את אבקתו. השלב הזכרי נמשך מספר שבועות והאבקה מוסיפה להיות מפוזרת מהפרח אפילו לאחר נשירת עלי הכותרת והבשלת הפירות. ברוב האוכלוסיות אין די פרחים בשלבים מתאימים כדי לאפשר האבקה-עצמית בין פרחים בשלב הנקבי לפרחים בשלב הזכרי של אותו פרט. עם זאת, נתקלתי במספר אוכלוסיות שלהן פקעות ענפות בעלות מספר התפצלויות אשר בהן כל ניצן מתפתח לענף קצר תת-קרקעי הנושא פרח בודד. במקרה זה יש לפרט הכלנית די פרחים לאפשר האבקה-עצמית שכן כאשר הפרח השלישי לדוגמא מתחיל לפרוח עם שלב מיני נקבי – הפרח הראשון הבוגר נמצא בשלב זכרי. טמפרטורות גבוהות מקצרות את הפער בין השלב הנקבי לשלב הזכרי, כך שהדיכוגמיה בצמחים בעלי פריחה מאוחרת הינה חלשה. פער זמנים זה מיטשטש גם בצמחי תרבות של הכלנית.
צמחי בר של הכלנית המצויה הינם הטרוזיגוטים באופן קיצוני, ובמאמצי ההכלאה שלי לטיפוח פרחים אף לאחר 12 דורות של הפריה עצמית נותרה הטרוזיגוטיות שאריתית. תחת תנאי הגידול המלאכותיים בהם גודלו עמדו רוב קווי הטיפוח הללו בהצלחה בזיווג השארים. אולם, במספר מקרים היו קלקולים: עקרות זכרית בקו אחד, ואובדן ייצור האנטוציאנין (לבקנות) בקווים אחרים, בהם ניסיתי להשיג טיפוח אמיתי לצבע פרח לילך. מאידך קיימת תבנית זיווג טובה בין פרטים שלהם תקופת פריחה שאינה מתואמת עם גל הפריחה העיקרי של האוכלוסיה: פריחה מוקדמת של פרחים שלהם עקרות-זכרית או פריחה מאוחרת של פרטים שלהם פרחים אדומים.
 תמונות ממיזם לייצור זרעי מכלוא של כלנית אשר הוקם במגדל עוז. נאספה אבקה מפרחים מסויימים של כלניות בר שפקעותיהן נתקבלו מאורה וגודלו בשטח פתוח במגדל עוז. באבקת פרחי בר אלה נעשה שימוש להאבקת פרחי כלניות תרבות מהזן 'דה-קאן' אשר פקעותיהן נקנו בחו"ל. בתמונות רואים את כלניות התרבות אשר גודלו בחממה. השקיות הלבנות הנראות בתמונה הן שקיות אשר נועדו לשמור על הזרעים מפיזור טבעי ©.
תמונות ממיזם לייצור זרעי מכלוא של כלנית אשר הוקם במגדל עוז. נאספה אבקה מפרחים מסויימים של כלניות בר שפקעותיהן נתקבלו מאורה וגודלו בשטח פתוח במגדל עוז. באבקת פרחי בר אלה נעשה שימוש להאבקת פרחי כלניות תרבות מהזן 'דה-קאן' אשר פקעותיהן נקנו בחו"ל. בתמונות רואים את כלניות התרבות אשר גודלו בחממה. השקיות הלבנות הנראות בתמונה הן שקיות אשר נועדו לשמור על הזרעים מפיזור טבעי ©.
האבקה
הצמחים מואבקים באמצעות רוח, ובמידה פחותה אף על ידי חרקים. המאביקים החשובים הן חיפושיות ממשפחת פרחיות הפרג Glaphyridae, האוכלות את האבקה ומשתמשות בפרחים כמחסה לילה ולהזדווגות , Kaesar et al. 2007)(Dafni et al. 1990). הפרחים נסגרים לעת ערב, נשארים סגורים כל הלילה ונפתחים לעת בוקר, מגיבים כנראה גם לכמות האור אך גם לטמפרטורה, תכונה הידועה בשם "תרמונסטיות" (Horovitz, 1975).
גופן של החיפושיות המבקרות בפרחי הכלנית מתכסה בגרגרי אבקה אותם הן מעבירות מפרחים בשלב הזכרי לפרחים בשלב הנקבי או הזכרי. חלק ממיני החיפושיות אף אוכלות את גרגרי האבקה. הפרחים אינם מייצרים צוף, אך הם מאוד אטרקטיביים ומושכים אליהם טווח נרחב של מיני חרקים התרים אחר אבקה. לצמחים הנמצאים בשלב הנקבי יש משיכה מעטה עבורם, אך, כמויות קטנות של גרגרי אבקה הנישאים ברוח או על ידי חיפושיות נלכדות בין הפפילות (גבשושיות) שעל פני העטיף הקטיפתי של הפרח וגורמים לביקורים נדירים של חרקים התרים אחרי מזון, וכתוצאה מכך, חרקים אלה גורמים גם להאבקה של פרחי הכלנית הנמצאים בשלב הנקבי (Horovitz 1991).
מדדנו את ההפצה של גרגרי אבקה של כלנית מצויה על-ידי הרוח ממוקד מרכזי אל סדרה של צלוחיות בעלות שקפים משומנים אשר הונחו על הקרקע. ניתוח התוצאות הראה כי המרחק הממוצע של הפצת גרגרי האבקה הוא שני מטרים, כאשר תבניות הפיזור נשלטות על-ידי כיוון הרוח ומערבולותיה. גרגר האבקה עצמו הינו חלק, ואם המשטחים עליהם הוא נשמר חלקים אף הם, הרי שנחיתה זאת היא רק הצעד הראשון בסדרה של העברות-חוזרות carry-overs. כלומר, מספר מעופים ונחיתות עד המיקום הסופי. על פי הערכה החיפושיות המבקרות בכלנית ונושאות אבקה, מבצעות מעופים של מספר מאות מטרים בין פרחים שונים. על כן בתכניות הטיפוח של הכלניות השתמשנו במרחקי בידוד של לפחות 500 מטרים.
התפוצה הרבה של אוכלוסיות הכלנית, שלעיתים תכופות הן גם רציפות, אשר בהן כל פרח מפיץ כמויות אדירות של גרגרי אבקה (כשני מיליון גרגרי אבקה לפרח), מהווה מקור חשוב של מזון עבור אוכלוסיית החרקים החורפיות דוגמת חיפושיות הפרג ודבורי-דבש אשר אינן מאביקות את הכלנית. "נדיבות" זאת מסתייעת בהאבקה באמצעות הרוח ומספקת שיעור לבוטנאים הנוטים לשקול שיקולי עלות (במקרה זה – תצוגה צבעונית וייצור אבקה) כנגד תועלת.
| הערות המערכת על האבקה רוח והאבקת חיפושיות בכלנית האדומה
אורה הורוויץ ז"ל, הייתה הראשונה אשר הוכיחה כי בכלנית המצויה חלה האבקת רוח בנוסף להאבקה חרקים. היא חשבה כי האבקת רוח היא הוקטור העיקר של תהליך ההפריה בכלנית ועל כן היה קשה לה להסביר את ההשקעה הגדולה של הצמח בעלי עטיף (כותרת בשפת העם) אדומים כה גדולים. אורה הסבירה את הערך האדפטיבי של קיום עלי העטיף בכך שפני העטיף מצויד ב"כוונה" בפפילות גבשושיות היוצרות מגע קטיפתי הלוכד את גרגירי האבקה החלקים הזעירים של הכלנית וזה מה שמתאים אותם להאבקת רוח ולא להאבקת חרקים! כלומר המשטח הגדול של העטיף הצבעוני הפתוח לרווחה מהווה שדה לכידה לגרגירי האבקה ומונע מהם להמשיך הלאה. איך יגיעו מהעטיף לצלקת במרכז הפרח? כפי שנכתב בפסקה לעיל, אורה לא האמינה ב"סיפור כמותי הסתברותי" ובנתה הסבר דטרמיניסטי של חיפושיות האוספות את גרגירי האבקה הנדירים הללו ומעבירות אותן לצלקות פרחים בשלב הנקבי. אורה הורוויץ ז"ל, לא האמינה כי חרקים המבקרים בעונת החורף בכלניות גורמים להאבקתן. על כך היו לנו (א.ש.) דיונים רבים בשנים בהם עבדנו באוכלוסיות כלנית מצויה אדומה בשפלה ובהרי-ירושלים (2005-2009) כאשר נוכחנו חזור ושנו כי דבורי-דבש, זבובים וחיפושיות מעבירים כמויות גדולות של אבקה מפרחים בשלב הזכרי לפרחים בשלב הנקבי. אורה האמינה אך מעט כי המכניזם הדרוויניסטי בטבע פועל באמצעות "כללים אגואיסטיים של עלות-תועלת" וסברה כי הכלניות מותאמות להאבקת רוח בלבד. זאת על אף הסתירות עם העובדות הקיימות (מדוע לכלנית פרח גדול צבעוני? ומדוע 27 מינים של חיפושיות פרחיות מבקרות בעיקר בפרחי צלחת אדומים ולא רק בכלניות?) אנו העדפנו להשאיר את הטקסט המקורי של אורה ומקווים שהקורא ייהנה מ"שני העולמות"… |
זרימת גנים פולימורפיזם בצבע האבקנים באוכלוסיות טבעיות של כלנית מצויה בישראל. מתוך עזבונה של א.הורוויץ ©.
פולימורפיזם בצבע האבקנים באוכלוסיות טבעיות של כלנית מצויה בישראל. מתוך עזבונה של א.הורוויץ ©.
האגוזיות הצמריות החד-זרעיות של פרי הכלנית נישאות למרחק עשרות מטרים על-ידי הרוח, ויש גם העברות חוזרות. לעיתים מצאתי גנוטיפים מיוחדים של כלנית אשר גידלתי בחוות הלימוד של האוניברסיטה בתלפיות במרחק שני קילומטרים מהחווה. אין ספק כי היו אלה צאצאים של הצמחים המיוחדים אשר גידלתי בחווה ולפיכך מרחק הפיזור האפשרי של זרעי הכלנית מגיע לפחות עד שני קילומטרים. אולם, מרחק ההפצה תלוי גם בטופוגרפיה, וחוות הלימוד שוכנת בשלוחה מוכת-רוחות בקו פרשת המים של הרי ירושלים. מאידך כל צאצאי המוטנטים החריגים של הכלניות בהם נתקלתי נשארו מתוחמים לצלע שלוחת החווה במשך דורות. זאת פרט ליוצא מן הכלל בו מצאתי מוטנט אחד בעמק מתחת לחווה. הוא הצליח לגדול ולפרוח בגן טיילת ארמון הנציב, כנראה בעקבות הרס עצום של בית -הגידול.
תודות
המחקר המולקולרי בוצע במעבדתה של דר’ נורית פירון, מהמכון לגידולי שדה במרכז וולקני, בהדרכתן של דר’ פירון ודר’ אורה הורוויץ.
ספרות
דפני א צהרי ח בן-שלמה ר ווריקן נ ונאמן ג 2020 צבע הפרחים, ההאבקה, והגנטיקה של הכלנית המצויה Anemone coronaria בישראל, כתב-עת "כלנית", מספר 7.הורוויץ א עברי י ולבנה מ 1988 "האדומים", טבע וארץ כרך ל' 6: עמוד 20–28.
הורוויץ א 2021 פס אדום בכלניות לבנות – תופעת טבע נדירה. כתב עת "כלנית" 8.
כוכבא ע 1998[ בהדרכת אורה הורוויץ] סקר השונות הגנטית המיטוכונדרית בכלנית המצויה ישראל. חיבור לשם קבלת תואר מוסמך במדעי הטבע. ירושלים, 1998
קדמן-זהבי א הורוויץ א 1980 תנאי יום ארוך מקדימים תרדמת כלניות. השדה, כרך ס"א (א’).
שמידע א 1981 – פרחי ראווה אדומים בצמחיה הים-תיכונית בישראל. "טבע וארץ", כ"ג:3 עמ' 118-126
שמידע א ודניאל ש 1994 – הנקבה הצנועה והזכר המתבלט: מדוע לכלנית המבוגרת יש טבעת לבנה? "טבע וארץ" חוברת 264 עמ' 20-23
——————–
Anikster Y and Feldman M 2019 Ora (Auguste) Horovitz (1920 – 2017) – In Memoriam, Is. J. of Plants Sciences.
Dafni, A., Bernhardt, P., Shmida A., Ivry, Y., Greenbaum, S., O’Tool, Ch., and Losito, L,. 1990 Red Bowl-shaped flowers: convergence for beetle pollination in the Mediterranean region. Isr. J. Bot. 39:81-92
Horovitz A and Zohary D 1966 Spontaneous variegation for perianth colour in wild Anemone coronaria. Heredity 21(3): 513-515.
Horovitz A and Zohary D 1971 Spontaneous Variegation for Perianth Colour in Wild Anemone coronaria, Heredity 21:513 – 515
Horovitz A Galil J and Zohari D 1975 Biological Flora of Israel 6: Anemone coronaria L. Isr. J. Bot. 24:26-41
Horovitz A 1976 Edaphic factors and flower colour distribution in the Anemoneae (Ranuculaceae). Plant Sys. Evo. 126: 239-242
Horovitz A and Harding J A 1978 Introduction of wild genes into commercial Anemone coronaria L. Final report to the USA-Israel Binational Science Foundation.
Horovitz A 1984 Seasonal pollen fertility in male sterile Anemone coronaria . Abs. Ann. Meet. Bot. Soc. Amer., J. Bot. 71: 5, Pt. 2): 98-99
Horovitz A 1985 Anemone coronaria and related species. In Abraham H. Halevy (Ed.) CRC Handbook of Flowering (I) CRC Press, Florida
Horovitz, A 1991 The pollination syndrome of Anemone coronaria L. : an insect-bias mutuakism. Acta Horticulturae 288: 283-287
Horovitz A 1995 The local gene pool of Anemone coronaria. Proc. 18th Eucarpia Symp. Sect. Ornamental Plant Improvement, classical and molecular approaches. Acta Horticulturaea 420:144-146
Keasar T Shmida A and Zylbertal A 2008 Pollination ecology of the red Anemone coronaria: honeybees may select for early flowering. Discussion Paper # 491. Center for the Study of Rationality: The Hebrew University of Jerusalem, Israel.
Keasar T Sabatinelli G and Shmida A 2010 Red anemone guild flowers as focal places for mating an feeding by Levant glaphyrid beetles. Biological Journal of the Linnaean Society 99: 808-817.
================
כל הזכויות שמורות ל"כלנית" ©
לציטוט: הורוויץ א 2021 פרופיל ביולוגי של הכלנית המצויה Anemone coronaria . כתב עת "כלנית" 8.
==========









